| |
Думай! Или "Супертренинг" без заблуждений
Вадим Протасенко
Введение
Я неслучайно остановил выбор названия этого
текста на том варианте, что написан чуть выше:
подготовленный читатель без труда узнает
в нём коллаж, составленный из заглавий двух
наиболее, пожалуй, популярных сегодня книг о
бодибилдинге. "Думай! Бодибилдинг без стероидов"
Стюарта Мак-Роберта и "Супертренинг" Майка
Менцера всколыхнули мир любительского спорта и
перевернули многие давно устоявшиеся
представления о теории тренировки. Точнее будет
выразиться так: Менцер впервые попытался создать
наконец хоть какую-то теорию. До него
большинство популярных книг и статей о
бодибилдинге представляли собой лишь
бессистемные наборы всевозможных (и зачастую
весьма противоречивых) тренировочных указаний,
а также каталоги упражнений с тяжестями. Менцер
же призвал рассматривать бодибилдинг как науку,
но почему-то выбрал в качестве
основы для этой науки не физиологию, а умозрительное
предположение о ключевой роли последнего
"отказного" повторения в механизме запуска роста
мышц. Как некогда Евклид создал свою геометрию,
опираясь на ряд аксиом о свойствах пространства,
так и Менцер создал свой "Супертренинг",
опираясь на аксиому об "отказном" повторении. У
Менцера это именно аксиома, поскольку он не
потрудился дать никакого обоснования своему
предположению. Но, как известно, помимо геометрии
Евклида существуют ещё и геометрии Лобачевского
и Римана, основанные на иных аксиомах, но столь
же внутренне непротиворечивые и логичные.
Вдохновлённый отменным стилем "Супертренинга" и
непоколебимой уверенностью его автора в своей
правоте и, следуя его советам, нарастив
10 килограммов "натуральных" мышц за полгода,
я стал ярым сторонником идей Менцера. Решив
найти физиологическое подтверждение его главной
аксиомы, я с головой окунулся в новые для себя
области познания — физиологию и биохимию
человека. Результат оказался для меня
неожиданным, но об этом чуть позже.
Ни для кого из любителей "железной игры", наверное,
не секрет, что теория тренировочного процесса
находится сегодня в довольно-таки неприглядном
состоянии. Все спортивные журналы полны статей с
многочисленными супермодными рекомендациями в
отношении тренинга, — но рекомендации эти,
увы, крайне противоречивы. "Движение с отягощением
должно быть мощным и взрывным", —
безапелляционно утверждают одни авторы. "К успеху
приводит только медленное подконтрольное движение",
— не менее решительно заявляют другие.
"Хочешь нарастить массу — работай с большими
весами", — дают уверенные указания третьи
специалисты. "Вес снаряда не имеет значения.
Главное — это техника и ощущение работы
мышцы", — противоречат им их оппоненты.
Тренироваться шесть дней в неделю утром и вечером
советует Арнольд Шварценеггер. Майк Менцер же
запрещает ученикам появляться в зале чаще двух
раз в неделю. Профессионалы бодибилдинга
расписывают для одного только бицепса комплексы
из шести упражнений. В свою очередь, Мак-Роберт
призывает вообще не тренировать руки
изолированными упражнениями. Пауэрлифтёры во
время своих циклов почти никогда не работают до
отказа, Менцер же уверяет, что работа не до отказа
— это зря потраченное время. А профи из
команды Джо Вейдера вообще советуют идти гораздо
дальше отказа с помощью форсированных повторений
и так называемого "стриптиза".
Это перечисление можно было бы ещё продолжать и
продолжать, но здесь обращает на себя внимание
не само обилие взаимоисключающих принципов
тренинга, а тот странный факт, что буквально у
каждого из этих принципов находятся сторонники,
сумевшие получить от его использования
положительный результат. Данный факт
способствовал распространению мнения, что
цельной, научной теории тренинга не может быть в
принципе. Я же утверждаю, что такая теория есть.
И терпеливый читатель вскоре сможет сам в этом
убедиться.
Слабым местом любой нынешней методики тренинга
является отсутствие у её авторов чёткого
представления о причинах роста мышц. Подавляющее
большинство специалистов даже не задумывается
над этим вопросом, остальные ограничиваются одной
лишь умозрительной идеей о тренировочном стрессе.
Полагая, что такое легкомыслие является бедой
только популярной спортивной литературы, я стал
искать ответ на вопрос о причинах роста мышц в
серьёзных научных трудах из области физиологии
мышечной деятельности. Однако меня постигло
разочарование: точными сведениями о том, что
именно происходит в мышце во время работы с
предельными нагрузками, современная наука, увы,
не располагает — или, во всяком случае, не
спешит поделиться этими сведениями с широким
кругом читателей. Связано это, на мой взгляд, в
первую очередь с тем, что основные исследования
физиологов всегда были направлены на обеспечение
потребностей, если можно так выразиться,
"классического" спорта, основой которого
является тренировка выносливости
(работоспособности). Максимум, что мне удалось
обнаружить — это исследование структурных
изменений в мышцах крыс после получасового
интенсивного плавания или бега в колесе. Понятно,
что по таким данным тяжело судить о реакции мышц
атлета на то напряжение, поддерживать которое
мышцы способны всего лишь несколько секунд.
Потеряв надежду найти готовые ответы, я решил
самостоятельно, путём анализа всех известных
мне сведений, смоделировать те процессы,
которые протекают в мышцах при нагрузках,
близких к предельным. Разработанная мной в
конце концов модель воздействия тренировки на
мышечный аппарат оказалась способной описать
практически все известные эмпирические факты
и позволила впервые найти ответы на ряд
вопросов, не получивших до настоящего момента
сколько-нибудь приемлемого объяснения со
стороны спортивных физиологов. Например, мне
удалось в общих чертах на молекулярном уровне
смоделировать механизм возникновения
микротравм мышечного волокна (источников
посттренировочной боли и общего стрессового
воздействия на организм), указать точные
условия их возникновения, а также найти
причины адаптации мышц к нагрузке и объяснить
феномен снижения посттренировочной боли при
регулярных тренировках. Ознакомиться с данной
информацией читатель сможет во второй части
текста, в коей, собственно, и изложена
суть моей теории. На основе этой теории в
третьей части текста я постарался объяснить,
почему работает "Супертренинг" Менцера,
тренировки в котором ограничиваются всего лишь
одним подходом в упражнении, и для чего
предназначены и как работают интервальные
(многоподходные) тренировки, а также в чём
секрет лифтёрского или штангистского силового
цикла, и вообще, чем определяется воздействие
большинства иных тренировочных методик и
приёмов. Для неподготовленного читателя
материал второй части статьи окажется, скорее
всего, слишком сложным для понимания, поэтому
я рекомендую внимательно изучить сначала
первую часть текста, в которой в более-менее
доступной форме изложены основные сведения о
строении и принципах функционирования мышц.
Должен заметить, что сведения, приведённые в
популярной литературе и даже в большинстве
учебных пособий по физиологии мышечной
деятельности для спортивных ВУЗов, с которыми
мне удалось ознакомиться, являются неполными.
Процессы синтеза белка клеткой в данной
литературе, как правило, даже не
рассматриваются. Судя по всему, именно по
этой причине очень многие люди, считающие себя
специалистами в области так называемой
"химии", на самом деле не имеют правильного
представления об анаболическом или, другими
словами, восстановительном механизме. Эти
"специалисты" любят рассуждать о "рецепторах"
стероидных гормонов, об их "забивке", о
"повышении чувствительности" оных рецепторов,
об образовании новых рецепторов и т.д., даже
не задумываясь о том, что эффект стероидных
гормонов реализуется через воздействие на
генетический аппарат, заключённый в ядре
клетки. Тем самым именно ядро и является
конечным "рецептором" тестостерона, кортизола
и ряда других гормонов. Не задумываются они и
о том, что важнейшее влияние на объём мышечной
ткани оказывает именно количество клеточных
ядер в мышце. Конечно, ядра мышечных клеток,
равно как и сама мышечная клетка, неспособны
к делению и размножению, но спортивные
физиологи как будто намеренно игнорируют
информацию о существовании в мышечных волокнах
так называемых "клеток-сателлитов"
(несформированных мышечных клеток),
сохраняющих способность к делению на
протяжении всей жизни человека —
благодаря чему и обеспечивается увеличение
количества мышечных ядер и регенерация
мышечных волокон при повреждениях мышечной
ткани. Как этот факт может повлиять на рост
объёма и силы мышц в результате тренировки,
читатель также сможет узнать из второй части
моего текста. Итак, повторяю, мне удалось
создать более-менее цельную теорию тренинга,
на физиологическом уровне объясняющую
(конечно, в общих чертах) воздействие
тренировки на мышечный аппарат человека и
позволяющую найти ответы на большинство
вопросов, связанных с улучшением характеристик
мышц.
Предвижу реакцию скептиков: как это человек
без специального образования посмел залезть в
самые дебри новой для себя науки, да ещё
набрался наглости выносить на суд публики
свои теории? Но что же ещё остаётся делать
нам, любителям "железной игры", если её
профессионалы не спешат предложить решение
волнующих нас проблем? Конечно, нам остаётся
полагаться только на собственные силы: в
конце концов, как писали классики, "спасение
утопающих — дело рук самих утопающих".
Часть I
Что нужно знать о
строении и принципе работы мышц
Различают три типа мышечной ткани: скелетную,
гладкую и сердечную. Функция сердечной ткани
понятна из названия, и её роль, я думаю,
объяснять не надо. О существовании гладких
мышц мы зачастую даже и не догадываемся,
поскольку это мышцы внутренних органов. И мы
лишены возможности напрямую управлять ими
(равно как, впрочем, и сердечной мышцей). Между
тем именно гладкие мышцы сужают просвет сосудов,
производят сокращение кишечника, способствуя
перемещению пищи, и выполняют ещё множество
других жизненно важных функций. В свою очередь,
задача скелетных мышц — перемещение частей
скелета друг относительно друга (отсюда и
название). Именно эти самые мышцы мы с таким
упорством и пытаемся нарастить на теле, и
именно их строение и свойства я буду
рассматривать в дальнейшем изложении.
Если заглянуть в
клетку
Как известно, все ткани организма имеют
клеточную структуру. Не представляют исключения
в этом плане и мышцы. Поэтому мне придётся
провести краткий экскурс в цитологию —
науку о клетке, и напомнить читателям о роли
и свойствах основных структур клетки. В грубом
приближении клетка состоит из двух важнейших
взаимосвязанных структур — цитоплазмы и
ядра.
Ядро содержит в себе молекулу ДНК, в которой
заключена вся наследственная информация. ДНК
— это полимер, закрученный двойной
спиралью. Каждая одинарная спираль ДНК состоит
из огромного количества четырёх видов
мономеров, называемых нуклеотидами.
Последовательность нуклеотидов в цепочке
кодирует все белки организма. Помимо хранения
всей наследственной информации, ядро
ответственно также и за размножение клетки
— деление. Деление клетки начинается с
разрывания двойной молекулы ДНК на две
отдельные цепочки нуклеотидов, каждая из
которых способна достроить себе пару из
набора нуклеотидов, находящихся в
свободном состоянии внутри клетки, чтобы тем
самым превратиться вновь в двойную
молекулу ДНК. Таким образом,
количество ДНК в ядре удваивается,
вслед за этим на две части делится ядро, а за
ним и вся остальная клетка.
Цитоплазма — это всё то, что в клетке
окружает ядро. Цитоплазма состоит из
цитозоли (клеточной жидкости), в которую
включены различные органеллы (части клетки),
такие, например, как митохондрии, лизосомы,
рибосомы и пр.
Митохондрии являются энергетическими
станциями клетки. В них с помощью различных
ферментов происходит окисление аминокислот,
углеводов и жирных кислот. Энергия,
выделяющаяся при этом окислении, идёт на
присоединение третьей фосфатной группы к
молекуле аденозиндифосфата (АДФ) с
образованием аденозинтрифосфата (АТФ) —
универсального источника энергии для всех
процессов, протекающих в клетке. Отсоединяя
третью фосфатную группу и вновь становясь
АДФ, АТФ выделяет запасённую ранее энергию.
Ферменты или энзимы — это вещества
белковой природы, в сотни и тысячи раз
увеличивающие скорость протекание химических
реакций. Практически все жизненно важные
химические процессы в организме происходят
только в присутствии специфических ферментов.
Лизосомы — это округлые пузырьки,
содержащие около пятидесяти ферментов.
Лизосомные ферменты расщепляют поглощённый
клеткой материал и собственные внутренние
структуры клетки (автолизис). Лизосомы,
сливаясь в фагосомы, способны переваривать
целые органеллы, подлежащие дезинтеграции.
Рибосомы — это органеллы, внутри которых
происходит сборка белковой молекулы.
Клеточная мембрана представляет собой просто
оболочку клетки. Мембрана обладает
избирательной проницаемостью, то есть
способностью пропускать одни вещества и
задерживать другие. Задача мембраны —
сохранять постоянство внутренней среды
клетки.
Строение мышцы
Структурно-функциональной единицей скелетной
мышцы является симпласт или мышечное волокно
— огромная клетка, имеющая форму
длинного цилиндра с заострёнными концами (в
дальнейшем под наименованиями "симпласт",
"мышечное волокно" и "мышечная клетка"
следует понимать один и тот же объект).
Длина мышечной клетки чаще всего
соответствует длине целой мышцы и
достигает 14 см, а диаметр равен нескольким
сотым долям миллиметра. Мышечное волокно, равно
как и любая другая клетка, имеет оболочку —
сарколему. Снаружи отдельные мышечные волокна
окружены рыхлой соединительной тканью, содержащей
кровеносные и лимфатические сосуды, а также
нервные волокна. Группы мышечных волокон
образуют пучки. Пучки, в свою очередь,
объединяются в целую мышцу, "упакованную" в
плотный чехол соединительной ткани, который
переходит на концах мышцы в сухожилия,
крепящиеся к костям. Усилие, вызываемое
сокращением мышечного волокна, передаётся через
сухожилия костям скелета и приводит их в
движение.
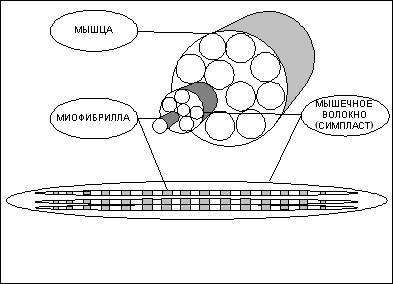 Рисунок 1
Управление сократительной активностью мышцы
осуществляется с помощью большого числа
мотонейронов (рис. 2) — нервных клеток,
тела которых лежат в спинном мозге, а
длинные ответвления, аксоны — в составе
двигательного нерва подходят к мышце. Войдя
в мышцу, аксон разветвляется на множество
веточек, и каждая из них подведена к
отдельному волокну. Таким образом, один
мотонейрон иннервирует целую группу волокон
(так называемую "нейромоторную единицу"),
которая работает как единое целое.
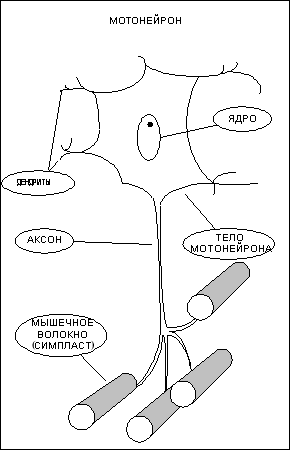 Рисунок 2
Мышца состоит из множества нейромоторных
единиц и способна работать не только всей
своей массой, но также и отдельными
пучками, что позволяет регулировать силу
и скорость сокращения. Для понимания
механизма сокращения мышцы необходимо
рассмотреть внутреннее строение мышечного
волокна.
От большинства других клеток оно отличается,
прежде всего, многоядерностью. Связано
это с особенностями формирования волокна при
развитии плода. Мышечное волокно образуется
на этапе эмбрионального развития организма
из клеток-предшественниц — миобластов.
Миобласты (неоформленные мышечные клетки)
интенсивно делятся, сливаются друг с другом
и образуют мышечную трубочку с центральным
расположением ядер. Затем в этой трубочке
начинается синтез миофибрилл —
сократительных элементов мышечного волокна
(см. ниже). Завершается формирование
мышечного волокна миграцией ядер на
периферию. Ядра мышечного волокна к этому
времени уже теряют способность к делению,
и за ними остаётся только функция хранения
информации для синтеза белка. Но не все
миобласты идут по пути слияния в мышечные
трубочки, часть из них обособляется в виде
клеток-сателлитов, располагающихся на
поверхности мышечного волокна, а именно в
сарколеме, между плазмолемой и базальной
мембраной — составными частями
сарколемы. Клетки-сателлиты, в отличие от
мышечных волокон, не утрачивают способности
к делению на протяжении всей жизни, что
обеспечивает увеличение общей массы
волокон и их обновление. Восстановление
мышечных волокон при повреждении мышцы
возможно только благодаря клеткам-сателлитам.
При гибели волокна скрывающиеся в его
оболочке клетки-сателлиты активизируются,
делятся и преобразуются в миобласты.
Миобласты сливаются друг с другом и образуют
новые мышечные трубочки, в которых затем
начинается сборка миофибрилл. То есть при
регенерации полностью повторяются события
эмбрионального (внутриутробного) развития
мышцы (Р.П.Женевская, Э.Г.Улумбеков и
Ю.А.Челышев)
Помимо многоядерности, отличительной чертой
мышечного волокна является наличие в
цитоплазме (у мышечного волокна её принято
называть саркоплазмой) тонких волоконец
— миофибрилл (рис. 1), расположенных
вдоль клетки параллельно друг другу. Число
миофибрилл в волокне достигает двух тысяч.
Миофибриллы являются сократительными
элементами мышечной клетки и обладают
способностью уменьшать свою длину при
поступлении нервного импульса, стягивая тем
самым мышечное волокно. Под микроскопом
видно, что миофибрилла имеет поперечную
исчерченность — чередующиеся тёмные
и светлые полосы. При сокращении миофибриллы
светлые участки уменьшают длину, а при
полном сокращении исчезают вовсе. Для
объяснения механизма сокращения миофибриллы
около пятидесяти лет назад Хью Хаксли
предложил модель скользящих нитей, которая
затем нашла подтверждение в экспериментах и
стала общепринятой.
Механизм сокращения волокна
Чередование светлых и тёмных полос в
миофибрильной нити определяется
упорядоченным расположением по длине
миофибриллы толстых нитей белка миозина и
тонких нитей белка актина; толстые нити
содержатся только в тёмных участках (А-диск)
(рис. 3), светлые же участки (I-диск)
не содержат толстых нитей. В середине I-диска
находится Z-линия, к ней крепятся тонкие
нити актина. Участок миофибриллы, состоящий
из А-диска (тёмной полосы) и двух
половинок I-дисков (светлых полос), называют
саркомером. Сокращение саркомера происходит
путём втягивания тонких нитей актина между
толстыми нитями миозина. Скольжение нитей
актина вдоль нитей миозина происходит
благодаря наличию у нитей миозина боковых
ответвлений, называемых мостиками. Головка
миозинового мостика сцепляется с актином и
изменяет угол наклона к оси нити, тем самым
как бы продвигая нити миозина и актина друг
относительно друга, затем головка
отцепляется, сцепляется вновь и вновь
совершает движение. Перемещение миозиновых
мостиков вдоль нитей актина можно сравнить с
гребками вёсел на галерах. Как перемещение
галеры в воде происходит благодаря движению
вёсел, так и скольжение нитей миозина
происходит благодаря гребковым движениям их
мостиков; существенное отличие состоит лишь
в том, что движение мостиков асинхронно.
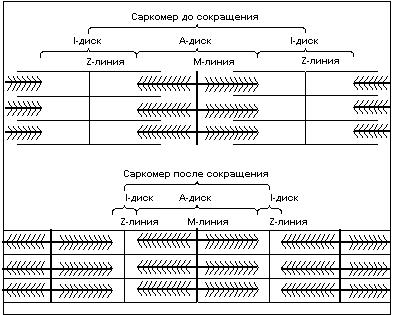 Рисунок 3
Тонкая нить представляет из себя
две спирально скрученные цепочки белка
актина. В канавках его спиральной цепочки
залегает двойная цепочка другого белка
— тропомиозина. В расслабленном
состоянии мостики миозина не имеют
возможности связаться с актином, поскольку
места́ его сцепления блокированы
тропомиозином. При поступлении же по аксону
двигательного мотонейрона нервного импульса
клеточная мембрана меняет полярность
заряда, и из специальных терминальных
цистерн, расположенных вокруг каждой
миофибриллы вдоль всей её длины, в
саркоплазму выбрасываются ионы
кальция (Ca++) (рис. 4).
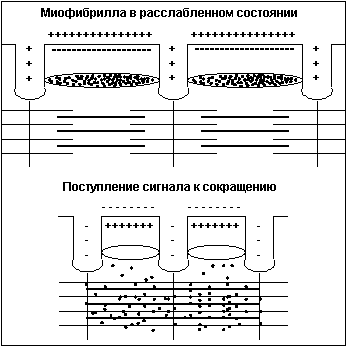 Рисунок 4
Под воздействием Са++ нить
тропомиозина входит глубже в канавку актина,
освобождая тем самым места́ для сцепления
миозина с актином, и мостики начинают цикл
гребков. Сразу после высвобождения иона
Са++ из терминальных цистерн он
начинает закачиваться обратно.
Соответственно, концентрация Са++
в саркоплазме очень быстро падает,
тропомиозин выдвигается обратно из канавки
и блокирует места сцепления миозиновых
мостиков. В результате мышечное волокно
расслабляется. Новый столь же короткий
нервный импульс опять выбрасывает
Са++ в саркоплазму и всё
повторяется. При достаточной частоте
импульсации (не менее 20 Гц) отдельные
мгновенные сокращения почти полностью
сливаются воедино, то есть достигается
состояние устойчивого сокращения,
называемое тетаническим сокращением или
состоянием гладкого тетануса.
Энергетика мышцы
Понятно, что для движения мостика требуется
энергия. Как я уже упоминал выше,
универсальным источником энергии в живом
организме является молекула АТФ. Под
действием фермента АТФазы АТФ гидролизуется,
отсоединяя фосфатную группу в виде
ортофосфорной
кислоты (Н3РО4), и
превращается в АДФ. При этом высвобождается
энергия.
АТФ + H2O = АДФ + H3PO4 + энергия
Головка миозинового мостика при контакте с
актином обладает АТФазной активностью —
то есть возможностью расщеплять АТФ и
получать энергию, необходимую для движения
вдоль нити актина. Запас молекул АТФ в мышце
ограничен, поэтому расходование энергии при
работе мышцы требуют постоянного восполнения
данного запаса. Мышца имеет четыре источника
воспроизводства энергии: расщепление
креатинфосфата, миокиназная реакция,
гликолиз и окисление органических веществ в
митохондриях.
Креатинфосфат обладает способностью
отсоединять от себя фосфатную группу и
превращаться в креатин, одновременно
присоединяя фосфатную группу к АДФ. Которая
тем самым превращается в АТФ.
креатинфосфат + АДФ = креатин + АТФ
Эта реакция получила название реакции
Ломана. Запасы креатинфосфата в волокне
невелики, поэтому он используется в
качестве источника энергии только на
начальном этапе работы мышцы, до момента
активизации других, более мощных
источников: гликолиза и кислородного
окисления. По окончании работы мышцы
реакция Ломана идёт в обратном направлении,
и запасы креатинфосфата в течение
нескольких минут восстанавливаются.
В условиях дефицита энергии при
значительном повышении концентрации АДФ
активируется миокиназная реакция, в ходе
которой фосфатная группа переносится с
одной молекулы АДФ на другую с
образованием АТФ и АМФ (аденозинмонофосфата).
АДФ + АДФ => АТФ + АМФ
Миокиназная реакция не играет
существенной роли в общем энергообмене
мышцы, однако она может быть источником
энергии в тех условиях, когда восполнение
энергии за счёт других источников
невозможно. Данная реакция, равно как и
креатинфосфатная, легко обратима.
Гликолиз — это процесс разложения
одной молекулы глюкозы (C6H12O6)
на две молекулы молочной кислоты
(C3H6O3) с
выделением энергии, достаточной для "зарядки"
двух молекул АТФ. Гликолиз протекает в
саркоплазме под воздействием десяти
специальных ферментов.
C6H12O6 + 2H3PO4 + 2АДФ = 2C3H6O3 + 2АТФ + 2H2O
Гликолиз идёт без потребления кислорода
(такие процессы называются анаэробными)
и способен быстро восстанавливать запасы
АТФ в мышце.
Окисление протекает в митохондриях под
воздействием специальных ферментов и
требует затрат кислорода, а соответственно,
и времени на его доставку. Такие процессы
называются аэробными. Окисление происходит
в несколько этапов: сначала идёт
гликолиз (см. выше), но образовавшиеся в
ходе промежуточного этапа этой реакции две
молекулы пирувата не преобразуются в
молекулы молочной кислоты, а проникают в
митохондрии, где окисляются в так
называемом "цикле Кребса" до углекислого
газа СО2 и воды Н2О
и дают энергию для производства ещё
тридцати шести молекул АТФ. Суммарное
уравнение реакции окисления глюкозы
выглядит так:
C6H12O6 + 6O2 + 38АДФ + 38H3PO4 = 6CO2 + 44H2О + 38АТФ
Таким образом, разложение глюкозы по аэробному
пути даёт энергию для восстановления тридцати
восьми молекул АТФ. То есть окисление в 19 раз
эффективнее гликолиза.
Типы мышечных волокон
В мышечных волокнах восполнение запасов энергии
идёт всеми способами одновременно. Но на
начальном этапе работы мышцы первостепенную
роль играет креатинфосфат. Креатинкиназная
реакция протекает почти мгновенно, скорость
воспроизводства энергии за счёт креатинфосфата
очень высока, поэтому мощные взрывные движения
реализуются в основном за счёт этого источника
энергии. Однако запасы креатинфосфата в
волокне очень невелики, их хватает лишь на
несколько секунд сокращения, поэтому при более
длительной работе на первый план выходят два
других более ёмких источника энергии —
гликолиз и окисление. Оба эти процесса
протекают в той или иной мере во всех
волокнах, но определённый тип волокон
специализируются, как правило, на каком-то
одном источнике. Поэтому волокна, в которых
доминирует гликолиз, принято называть
гликолитическими, а волокна, в которых
доминирует окисление, соответственно, —
окислительными. Но такое чёткое разделение
наблюдается не во всех волокнах — в
ряде волокон оба процесса активны в равной
мере. Такие волокна называются
окислительно-гликолитическими.
Разделение по способу получения энергии
неслучайно: дело в том, что у разных
источников энергии разная скорость её
воспроизводства и разная ёмкость. Так,
окисление требует времени на доставку
кислорода из крови в волокно, а затем в
митохондрии, где и происходит ресинтез АТФ,
которую затем ещё необходимо доставить к
работающим миозиновым мостикам. Гликолиз
же протекает непосредственно в саркоплазме
и восстанавливает запасы АТФ
значительно быстрее, чем окисление. Поэтому скорость
расходования энергии в окислительных волокнах
заметно ниже, чем в гликолитических.
Скорость расходования энергии, а
соответственно, сила и скорость сокращения
волокна, определяются типом его миозина.
Изоформа миозина, обеспечивающая высокую
скорость сокращения мышцы, — быстрый миозин
— характеризуется высокой активностью
АТФазы и, соответственно, высокой скоростью
расходования АТФ. Изоформа миозина с меньшей
скоростью сокращения — медленный
миозин — характеризуется меньшей
активностью АТФазы. Волокна с высокой
активностью АТФазы и скоростью
расходования АТФ принято называть
быстрыми волокнами; волокна, характеризующиеся низкой
активностью АТФазы и меньшей скоростью
расходования АТФ, — медленными волокнами.
Описанная выше взаимосвязь между
скоростью сокращения волокон и их
энергетическими механизмами позволяет
констатировать, что быстрые волокна по
способу получения энергии являются
гликолитическими, а медленные —
окислительными. Волокна промежуточного
типа — окислительно-гликолитические
— обычно относятся к быстрому типу,
так как при необходимости они могут
развивать значительное усилие и высокую
скорость сокращения, используя гликолиз
в качестве основного источника энергии,
в то же время при низкой интенсивности
сокращения для воспроизводства энергии
ещё достаточно эффективно используют
окисление.
Хотя окисление воспроизводит энергию
медленнее, чем гликолиз, оно зато куда
более эффективно в использовании
органического топлива: распад одной
молекулы глюкозы по гликолитическому пути
даёт энергию для зарядки всего лишь двух
молекул АТФ, в то время как окисление
глюкозы заряжает энергией сразу 38
молекул АТФ. Кроме того, молочная
кислота, возникающая при гликолизе и
накапливающаяся в мышечном волокне, вносит
существенный вклад в развитие мышечного
утомления. Все перечисленные обстоятельства
приводят к тому, что медленные волокна способны
на более длительную работу, чем быстрые. Так,
быстрые волокна утомляются при работе от
десятков секунд до нескольких минут, в то
время как медленные волокна способны
работать часами.
Для доставки кислорода от капилляров крови
вглубь мышечного волокна используется белок
миоглобин, имеющий красный цвет.
Окислительные волокна содержат этого белка
гораздо больше, чем гликолитические,
которым кислород нужен в меньшей степени,
поэтому под микроскопом окислительные
волокна имеют выраженный красный цвет.
Гликолитические же волокна выглядят гораздо
бледнее. Отсюда за обоими данными типами
волокон закрепилось ещё одно название:
красные и белые.
Тип мышечного волокна зависит от
иннервирующего его мотонейрона. Все волокна
одного мотонейрона принадлежат к одному
типу. Интересный факт: при подключении к
быстрому волокну аксона медленного
мотонейрона это быстрое волокно
перерождается, то есть меняет свой тип —
и наоборот. До недавнего времени
существовало две точки зрения на зависимость
типа волокна от типа мотонейрона. Одни
исследователи полагали, что свойства волокна
определяются частотой импульсации
мотонейрона; другие — что данные
свойства определяются влиянием
гормоноподобных веществ, поступающих из
нерва (вещества эти до настоящего времени
так и не выделены). Исследования последних
лет показали, что право на существование
имеют обе приведённые точки зрения, то есть
воздействие мотонейрона на волокно
осуществляется обоими способами.
Регуляция силы и скорости мышечного сокращения
Как известно, человек обладает способностью
произвольно регулировать силу и скорость
мышечного сокращения. Реализуется эта
способность несколькими путями. Об одном из
них я уже писал — это управление
частотой нервной импульсации. Подавая
волокну одиночные команды на сокращение,
можно добиться лёгкого его напряжения.
Например, поддерживающие позу мышцы слегка
напряжены даже тогда, когда человек отдыхает.
Повышая частоту импульсов, можно увеличивать
силу сокращения до максимально возможной для
данного волокна в данных условиях работы
— которая достигается при слиянии
отдельных коротких импульсов-всплесков в
сплошной гладкий тетанус. Сила, развиваемая
волокном в состоянии тетануса, не всегда
одинакова — она очень зависит от
скорости движения. При статическом
напряжении (когда длина волокна остаётся
постоянной) сила, развиваемая волокном,
всегда бывает больше, чем при сокращении
волокна. И чем быстрее сокращается волокно,
тем меньшую силу оно способно развить.
Максимальную же силу, способность
противостоять нагрузке, волокно развивает
в условиях негативного движения, то есть
при удлинении волокна.
При отсутствии нагрузки волокно сокращается
с максимальной скоростью. При увеличении
нагрузки скорость сокращения волокна
уменьшается и по достижении определённого
уровня нагрузки падает до нуля; при
дальнейшем увеличении нагрузки волокно уже
удлиняется. Причину в различии силы волокна
при различных направлениях движения легко
понять, рассмотрев уже приведённый ранее
пример с гребцами и вёслами.
Дело в том, что после завершения "гребка"
миозиновый мостик некоторое время ещё
находится в состоянии сцепления с нитью
актина (весло после гребка тоже не сразу
извлекается из воды, а находится
погружённым ещё некоторое время). В том
случае, когда гребцы плывут вперёд
(позитивное движение), вёсла, остающиеся
погружёнными в воду ещё и после завершения
гребка, тормозят движение и мешают плыть.
В то же время если лодка буксируется
назад, а гребцы сопротивляются этому
движению, то погружённые вёсла опять-таки
мешают движению, и буксиру приходится
прилагать дополнительные усилия. То есть
при сокращении миофибриллы сцепленные
мостики мешают этому сокращению и тем
самым уменьшают силу волокна; при
негативном же движении (при удлинении
миофибриллы) не успевшие отцепиться
мостики также мешают движению — но в
этом случае они как бы поддерживают
опускающийся вес, что и позволяет волокну
развивать значительно большую силу. Легче
всего понять различия между статическим
напряжением, позитивным и негативным
движениями можно, взглянув на рисунок 5.
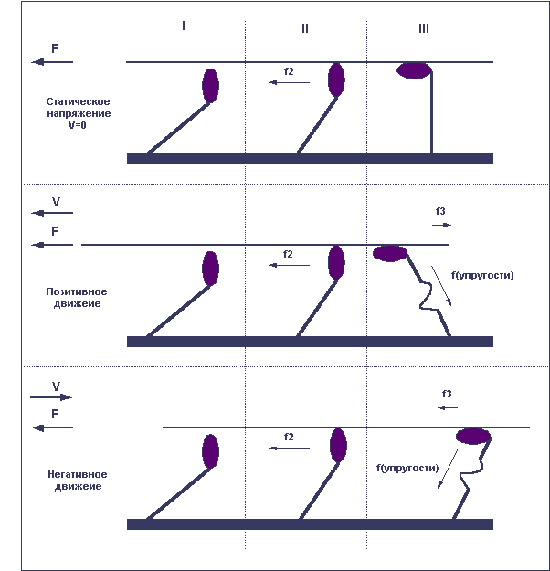 Рисунок 5
Итак, здесь было рассказано об основных
факторах, влияющих на силу и скорость
сокращения отдельного волокна. Что же
касается силы сокращения целой мышцы, то
она, понятно, зависит от количества волокон,
вовлечённых на данный момент в работу.
Вовлечение волокон в работу
При поступлении от центральной нервной
системы (ЦНС) к расположенным в спинном
мозге мотонейронам возбуждающего сигнала
(запускающего импульса) мембрана
мотонейрона поляризуется, и он генерирует
серию коротких импульсов, направляемых по
аксону к мышечным волокнам. Чем сильнее
воздействие на мотонейрон (чем сильнее
поляризация мембраны), тем выше частота
генерируемого им импульса — от
небольшой стартовой (4-5 Гц), до
максимально возможной для данного
мотонейрона частоты (50 Гц и более).
Быстрые мотонейроны способны генерировать
гораздо более высокочастотный импульс,
чем медленные, поэтому сила сокращения
быстрых волокон гораздо больше подвержена
частотной регуляции, чем сила медленных.
В то же время имеется и обратная связь
мотонейронов с мышцей, от которой поступают
тормозящие сигналы, уменьшающие поляризацию
мембраны мотонейрона и снижающие частоту
его импульсов. Каждый мотонейрон имеет свой
порог возбудимости. Если разность между
возбуждающими и тормозящими сигналами
превышает этот порог, и на мембране
достигается необходимый уровень поляризации,
то мотонейрон вовлекается в работу. Медленные
мотонейроны имеют, как правило, низкий порог
возбудимости, а быстрые — высокий.
Мотонейроны же целой мышцы имеют широкий
спектр значений этого параметра. Таким
образом, при повышении силы сигнала ЦНС
активируется всё большее число мотонейронов,
а мотонейроны с низким порогом возбудимости
увеличивают частоту генерируемого импульса.
Когда требуется лёгкое усилие (например,
при ходьбе или при беге трусцой), то
активируется необходимое число медленных
мотонейронов и соответствующее количество
медленных волокон. Ввиду высокой
выносливости этих волокон такая работа может
поддерживаться очень долго. По мере же
увеличения нагрузки ЦНС приходится посылать
всё более и более сильные сигналы, и в
работу вовлекается всё большее и большее
число мотонейронов (а следовательно, и
мышечных волокон). Одновременно те волокна,
что уже работали, увеличивают силу сокращения
по причине увеличения частоты импульсации,
поступающей от мотонейронов. По мере
увеличения нагрузки в работу включаются
окислительно-гликолитические волокна, но по
достижении определённого уровня нагрузки
— 20-25% от максимума (например, во
время подъёма в гору или финального спурта)
— силы окислительных волокон
становится недостаточно. И тогда посылаемый
ЦНС сигнал ещё больше усиливается и
включает в работу быстрые —
гликолитические — волокна. Быстрые
волокна значительно повышают силу сокращения
мышцы, но зато, обратным образом, быстро
утомляются, и потому в работу начинает
вовлекаться всё большее и большее их
количество. И если уровень внешней нагрузки
не уменьшается, то работу в скором времени
приходится останавливать из-за усталости.
Скорость сокращения мышцы зависит от
соотношения внешней нагрузки и количества
вовлечённых в сокращение волокон. Мобилизация
относительно небольшого количества волокон
сперва приводит к медленному движению —
стоит только совместной силе сокращающихся
волокон превысить уровень сопротивления
нагрузки. Мобилизация большого количества
волокон при той же внешней нагрузке приводит
к увеличению скорости сокращения, а
увеличение внешней нагрузки или падение силы
волокон в результате усталости (при
неизменном количестве мобилизованных волокон)
приводят к падению скорости сокращения. При
предельных нагрузках — например, при
подъёме максимального веса или подъёме
относительно небольшого веса, но с
максимальной скоростью — сокращается
сразу максимально возможное для данного
индивида число волокон.
По вопросу вовлечения волокон в работу при
предельных нагрузках в литературе приведены
противоречивые сведения. Так, Х.Тюннеманн и
Ю.Хартманн утверждали, что даже у
тренированных спортсменов при предельных
нагрузках активно не более 85% от общего
количества двигательных единиц в мышце. С
другой стороны, Гурфинкель и Левин привели
данные, что большая часть двигательных
единиц рекрутируется уже при нагрузках
до 50% от максимума, и в дальнейшем, при
повышении силы, к работе подключается лишь
небольшая часть (около 10%) самых крупных
двигательных единиц. Так что рост силы
от 75% до 100% происходит уже не за счёт
вовлечения новых двигательных единиц, а за
счёт повышения частоты импульса,
генерируемого мотонейронами.
Независимо от того, чья точка зрения
является правильной, важно одно: сила мышцы
зависит от силы импульса ЦНС, а то, чем на
самом деле обусловлен рост силы при
нагрузках свыше 75% от максимальной (только
ли повышением частоты, генерируемой
мотонейронами, или же ещё и вовлечением в
работу новых мотонейронов) — не
принципиально. Важно обратить внимание
также и на то, что в мышцу встроен
ограничительный механизм контроля за
развиваемым напряжением. Этот контроль
осуществляется через сухожильные органы
Гольджи. Сухожильные рецепторы регистрируют
напряжение и при превышении критического
значения оказывают тормозящее воздействие
на мотонейроны. Считается, что именно
отключение контроля за напряжением и
является разгадкой проявления той
"сверхсилы" человека, которая
регистрируется в экстремальных ситуациях.
Часть II. Теория тренировки
Вступление
Первую часть данного текста я посвятил
краткому изложению основ физиологии
мышечной деятельности, и теперь пришла
пора ознакомить читателей с основами
классической теории тренировки.
Организм — это саморегулируемая
система, стремящаяся к поддержанию
постоянства внутренней среды. Физическая
нагрузка оказывает выраженное воздействие
на внутреннюю среду мышц и организма в
целом, смещая многие биохимические
показатели от уровня, характерного для
состояния покоя, к уровню, соответствующему
состоянию деятельности. Степень этих
изменений зависит от характера и
интенсивности физической нагрузки и
индивидуальной реакции на неё организма,
отражающей уровень тренированности. Сразу
после прекращения нагрузки в организме
начинаются процессы, которые стремятся
восстановить исходное состояние,
соответствующее гомеостазу покоя. В ходе
этих процессов закрепляются изменения,
позволяющие в дальнейшем минимизировать
возмущение внутренней среды при
аналогичных нагрузках.
Спортивную тренировку следует рассматривать
как процесс направленного приспособления
организма (адаптации) к воздействию
тренировочных нагрузок.
Различают срочную и долговременную
адаптации. Срочная адаптация — это
ответ организма на однократное
воздействие тренировочной нагрузки,
выражающийся в "аварийном" приспособлении
к изменившемуся состоянию своей внутренней
среды. Ответ этот сводится преимущественно
к изменениям в энергетическом обмене и к
активации высших нервных центров,
ответственных за регуляцию энергетического
обмена. Что же касается долговременной
адаптации, то она формируется постепенно
на основе многократной реализации срочной
адаптации путём суммирования следов
повторяющихся нагрузок.
В протекании процессов адаптации можно
различить специфическую компоненту и
общую адаптационную реакцию. Процессы
специфической адаптации затрагивают
внутриклеточный энергетический и
пластический обмен и связанные с ним
функции вегетативного обслуживания,
которые специфически реагируют на
данный вид воздействия сообразно его
силе.
Общая адаптационная реакция развивается
в ответ на самые разные раздражители
(независимо от их природы) в том случае,
если сила этих раздражителей превышает
некий пороговый уровень. Реализуется
общая адаптационная реакция благодаря
возбуждению симпато-адреалиновой и
гипофизарно-адренокортикальной систем.
В результате их активации в крови и
тканях повышается содержание
катехоламинов и глюкокортикоидов, что
способствует мобилизации энергетических
и пластических резервов организма. Такая
неспецифическая реакция на раздражение
была названа "синдром стресса", а
раздражители, вызывающие эту реакцию,
получили название "стресс-факторы".
Общий адаптационный синдром сам по себе
не является основой адаптации к
тренировочным нагрузкам, он лишь призван
на системном уровне обеспечивать
протекание специфических адаптационных
реакций, которые и формируют
приспособление организма к конкретным
видам нагрузки.
Несмотря на различную природу процессов
специфической адаптации, можно выделить
общие закономерности их протекания.
Основу специфической адаптации составляют
процессы восстановления растраченных во
время мышечной работы энергетических
ресурсов, разрушенных структур клеток,
смещённого водно-электролитического
баланса и др. Наглядно проследить
закономерности протекания восстановительных
процессов можно на примере восстановления
энергетических ресурсов организма,
поскольку при физических нагрузках наиболее
выраженные изменения обнаруживаются именно
в сфере энергетического обмена.
Изменения в энергетическом обмене
Мышечная работа в зависимости от её
интенсивности и длительности приводит к
снижению уровня креатинфосфата в мышцах, а
также к истощению запасов внутримышечного
гликогена, гликогена печени и резервов
жиров. Интенсивно протекающие после
прекращения нагрузки процессы
восстановления приводят к тому, что в
определённый момент отдыха после работы
уровень энергетических веществ начинает
превышать исходный "дорабочий" уровень. Это
явление получило названия "сверхвосстановление"
или "суперкомпенсация" (рис. 1).
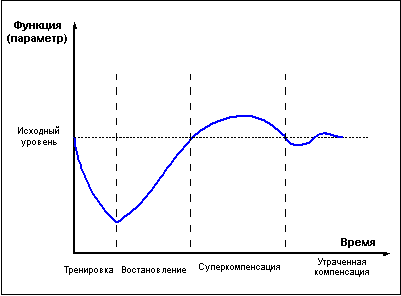
Рис. 1 |
Фаза суперкомпенсации длится не вечно,
уровень запасов энергетических веществ
постепенно возвращается к норме, испытывая
некоторые колебания возле состояния
равновесия. Чем больше был расход энергии
при работе, тем интенсивнее идёт
восстановление и тем значительнее
оказывается превышение исходного уровня в
фазе суперкомпенсации. Однако это правило
применимо лишь до какого-то предела.
При истощающих нагрузках, приводящих к накоплению
слишком большого количества продуктов распада,
скорость восстановительных процессов
уменьшается, фаза суперкомпенсации
откладывается и оказывается выраженной в
меньшей степени.
Похожим образом идёт восстановление не только
энергетических, но и пластических ресурсов
организма, и даже целых тренируемых функций.
Напряжение в ходе физической нагрузки систем,
ответственных за реализацию той или иной
функции, сначала приводит к снижению
функциональных возможностей организма, но
затем во время отдыха достигается состояние
суперкомпенсации тренируемой функции, длящееся
определённое время, а ещё через какое-то
время, при условии отсутствия повторных
нагрузок, уровень тренируемой функции вновь
снижается, — то есть наступает фаза
утраченной суперкомпенсации (рис. 1).
Выработка долговременной адаптации
становится возможной только в том случае, если
тренировки ведутся по определённым правилам,
благодаря чему их эффекты суммируются.
Проведение повторных тренировок в фазе
утраченной суперкомпенсации (слишком редкие
тренировки) (рис. 2) не сможет привести к
закреплению тренировочного эффекта, поскольку
каждая последующая тренировка проводится
после возвращения функциональных возможностей
организма к исходному уровню.
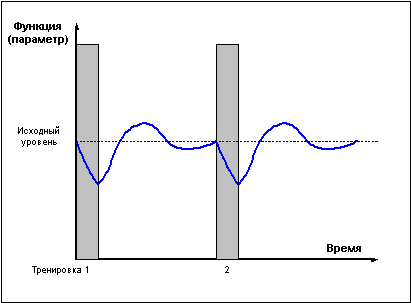
Рис. 2 |
В свою очередь, слишком частые тренировки,
прерывающие стадию восстановления до
достижения эффекта суперкомпенсации (рис. 3)
приводят к отрицательному взаимодействию
тренировочных эффектов и к снижению
функциональных возможностей организма.
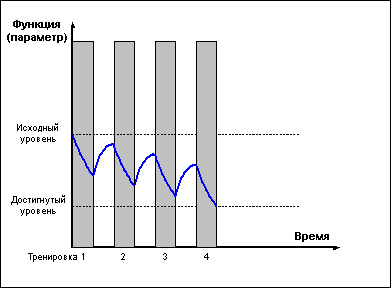
Рис. 3 |
И только проведение повторных тренировок в
фазе суперкомпенсации (рис. 4) приводит к
положительному взаимодействию тренировочных
эффектов, закреплению следов срочной
адаптации, росту тренируемой функции и
формированию долговременной адаптации.
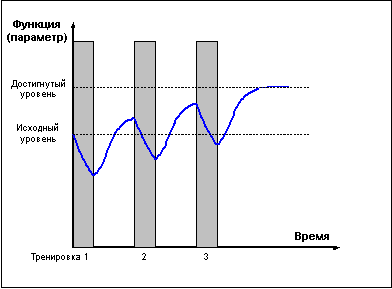
Рис. 4 |
Однако приведённые выше правила не следует
воспринимать узко, однолинейно. Требование
задавать нагрузку только на стадии
суперкомпенсации справедливо лишь в
долгосрочной перспективе. В рамках же одного
тренировочного микроцикла возможны и серии
тренировок в стадии недовосстановления (рис. 5),
приводящие к более глубокому истощению
тренируемой функции — что может быть
использовано затем либо для получения более
мощного роста функциональных возможностей в
стадии суперкомпенсации, либо для вызванного
тактической необходимостью переноса во
времени эффекта суперкомпенсации.
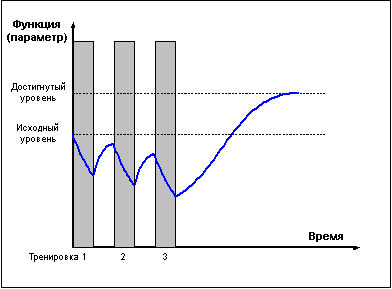
Рис. 5 |
На первый взгляд может показаться, что
составление эффективных тренировочных
программ является делом несложным: тут, мол,
достаточно лишь определить уровень нагрузки,
необходимый для достижения максимальной
суперкомпенсации той или иной функции, а
также время наступления фазы
суперкомпенсации — и тогда можно будет
задавать повторные нагрузки с необходимой
частотой, постоянно получая положительную
сумму тренировочных эффектов. Однако на
практике данный принцип построения
тренировки реализовать в полной мере почти
никогда не удаётся.
Всё дело в том, что различные параметры и
функции, вносящие вклад в общую
тренированность (необходимую для того или
иного вида спорта), имеют не только разное
время восстановления и достижения
суперкомпенсации, но также и разную
длительность фазы суперкомпенсации. Так,
фаза суперкомпенсации уровня креатинфосфата
в мышце достигается всего через несколько
минут отдыха после нагрузки, приводящей к
существенному снижению его уровня. Для
достижения же выраженной суперкомпенсации
запасов гликогена в мышцах требуется не
менее двух-трёх суток, и к этому моменту
уровень креатинфосфата уже вступает в фазу
утраченной суперкомпенсации. В свою
очередь, для восстановления структур клеток,
разрушенных в ходе тренировок, требуется
ещё большее время, в течение которого
уровень гликогена в мышцах уже может
вернуться к исходному уровню. Так что
заявления некоторых "гуру" бодибилдинга о
том, что время восстановления мышцы после
тренировки должно составлять N часов (дней),
без указания на то, о восстановлении какой
именно ведущей функции идёт речь, являются,
мягко выражаясь, сомнительными. Задать такой
период отдыха между тренировками, который
позволил бы получать усиление всех
тренируемых функций одновременно — дело
просто невозможное.
Поэтому в классическом подходе к спортивной
тренировке годичный (и даже многолетний)
период тренировок разбивают на микро- и
макроциклы, в ходе которых ставятся задачи
по повышению определённых способностей.
Чередование тренировочных занятий в ходе
микроциклов осуществляется таким образом,
чтобы физические нагрузки, направленные на
наращивание определённой двигательной
способности, задавались через промежутки
времени, обеспечивающие суперкомпенсацию
ведущей функции, а нагрузки иной
направленности, применяемые в этот период,
не оказывали отрицательного влияния на
восстановление основной функции.
Однако такой метод срабатывает только при
тренировке, если можно так выразиться,
"взаимонезависимых" функций или параметров.
Если же какая-то двигательная способность
зависит от развития сразу нескольких
функций или параметров, испытывающих
напряжение в ходе одного тренировочного
занятия и имеющих разное время
восстановления, то в течение микроцикла
приходиться варьировать интенсивность и
объём тренировок, накладывая волны
восстановления различных параметров друг на
друга таким образом, чтобы получить
суперкомпенсацию основных тренируемых
функций к моменту завершения микроцикла.
На рисунке 6 представлен простейший вариант
построения микроцикла для двух тренируемых
функций, имеющих разное время
восстановления. В течение микроцикла одна
функция испытывает последовательное
положительное суммирование тренировочных
эффектов, в то время как другая функция
последовательно вводится в стадию истощения
и достигает суперкомпенсации лишь во время
отдыха либо снижения нагрузки к концу
микроцикла. Реальная картина тренировок,
конечно, куда сложнее, поскольку число
тренируемых параметров и функций обычно не
ограничивается двумя, а доходит до десятка.
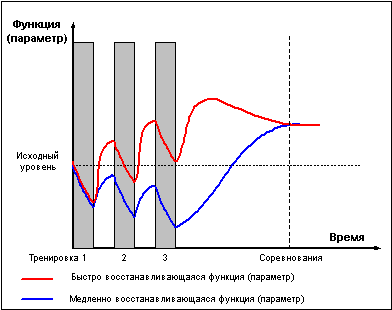
Рис. 6 |
Так уж сложилось, что классическая теория
тренировки основывается на изучение
процессов, приводящих к росту
работоспособности мышц и организма в самых
различных режимах работы. Дело в том, что
основой практически всех видов спорта
является именно работоспособность, а потому
традиционная цель планируемых адаптационных
изменений в организме спортсмена —
выход на новый уровень работоспособности.
Гипертрофия же мышц в классическом спорте
никогда не была целью тренинга и потому
воспринимается лишь как побочный продукт
развития основных двигательных качеств.
Более того, гипертрофия мышц иногда может
даже мешать достижению стоящих перед
спортсменом целей. А вот посетителей
тренажёрных залов, за редким исключением,
интересует не столько повышение
двигательных способностей, сколько именно
гипертрофия мышц — хотя, безусловно,
увеличение выносливости мышц способствует
гипертрофии мышечной ткани. За счёт чего?
Во-первых, за счёт того, что процессы,
направленные на улучшение доставки
кислорода к мышцам, существенно развивают
капиллярную сеть. Во-вторых, за счёт того,
что тренировка окислительной активности
мышц приводит к значительному росту в
саркоплазме мышечных волокон количества и
объёма митохондрий — энергетических
станций клетки. В-третьих, за счёт того,
что суммирующиеся процессы суперкомпенсации
внутримышечных запасов гликогена приводят
к заметному его накоплению, что, в свою
очередь, увеличивает объём саркоплазмы
мышечного волокна. К гипертрофии мышц
приводит накопление и иных веществ,
ответственных за энергообеспечение мышечной
деятельности — таких, например, как
креатинфосфат — что также увеличивает
объём саркоплазмы, и даже не столько за
счёт роста объёмов самих этих веществ,
сколько за счёт сопутствующего увеличения
объёма внутриклеточной жидкости. Таким
образом, тренировка работоспособности мышц
приводит к гипертрофии мышечных волокон в
первую очередь за счёт увеличения объёма
саркоплазмы.
Однако наибольший вклад в рост объёмов и
силы сокращения мышечных волокон вносит
гипертрофия именно самих миофибрилл, а все
остальные компоненты мышечных клеток
призваны лишь обеспечивать их, миофибрилл,
сократительную активность. Как размер
топливных баков самолёта зависит от
мощности, от "аппетита" его турбин, так и
объём саркоплазмы мышечного волокна зависит
от параметров сократительного аппарата
клетки (ну и, конечно, от объёма работы,
регулярно выполняемой мышцей).
Поскольку миофибриллы представляют собой
белковые нити, увеличение количества и
поперечного сечения миофибрилл в мышечном
волокне напрямую связано с интенсивностью
синтеза белка. То, что тренировка
интенсифицирует синтез белка — это
сегодня неопровержимый факт. Но вот вопрос:
как и почему это происходит?
Как строится белок
Молекула белка представляет собой цепочку
аминокислот, число которых колеблется от
нескольких десятков до нескольких десятков
тысяч. Всего в природе насчитывается
более трёхсот видов аминокислот, но для
строительства белка живые существа
использует только двадцать две. Свойства
белка определяются последовательностью
аминокислот в цепочке, а также
пространственной конфигурацией самой
цепочки (так называемыми "вторичной" и
"третичной" структурами белка).
Строительство молекул белка происходит как
из аминокислот, поступающих в организм с
белковой пищей, так и из аминокислот,
синтезируемых самим организмом. Упрощённо
процесс синтеза белка изображён на рисунке 7.
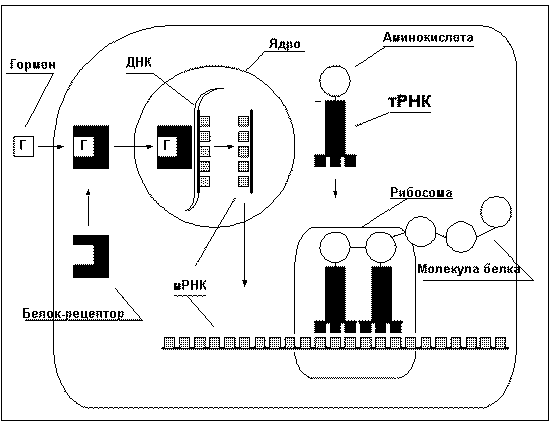
Рис. 7 |
Программы строительства всех белков
организма записаны в ДНК ядра
клетки и её частичных копиях — РНК
— в виде последовательности нуклеотидов. Каждая
аминокислота записывается определённой
комбинацией трёх нуклеотидов, называемой
"кодон". Последовательность кодонов в ДНК
и РНК определяет последовательность
аминокислот в белке. Последовательность
нуклеотидов, которая кодирует один белок,
называется "ген". Эта последовательность
считывается с ДНК и записывается в
матричной РНК (мРНК); процесс этот носит
название "транскрипция". мРНК — это
как бы кусочек ДНК, способный выходить из
ядра в протоплазму, где он закрепляется
на рибосомах. Транспортные РНК (тРНК)
доставляют к мРНК аминокислоты. Один конец
тРНК узнаёт на мРНК соответствующий
кодон и прикрепляется к нему. Аминокислота,
находящаяся на другом конце тРНК, сцепляется
с аминокислотой соседней тРНК; таким
образом выстраивается цепочка белка.
Синтез белка — это очень сложный
процесс, и его интенсивность зависит от
огромного числа факторов. Транскрипция
мРНК в ядре клетки начинается под
воздействием стероидных гормонов,
вырабатываемых железами внутренней
секреции и переносимыми к клеткам кровью.
Проникнув из крови внутрь клеток, гормоны
с помощью белков-рецепторов доставляются
в ядра и разблокируют участки цепочек
ДНК, ответственные за тот или иной белок,
после чего, собственно, транскрипция мРНК
и становится возможной. Для запуска
транскрипции РНК необходимо также
развернуть спираль ДНК, распрямить её,
для чего используется фермент
РНК-полимераза.
Сильнейшее влияние на синтез белка
оказывает гормон роста (СТГ или
соматотропин). По химическому составу
соматотропин сам является белком, поэтому
он не может свободно проникать в клетку
(в отличие от стероидных гормонов) —
он лишь воздействует на рецепторы,
расположенные на поверхности клетки.
Механизм действия гормона роста до конца
не изучен, но уже точно известно, что он
стимулирует деятельность РНК-полимераз и
рибосомного аппарата клетки.
Помимо всего прочего, для сборки белка
требуется наличие в клетке достаточного
количества аминокислот и запасов энергии.
Без аминокислот не из чего будет строить
белок, а энергия нужна для подпитывания
механизма сборки белковой молекулы.
Итак, для успешного синтеза белка
требуются, как минимум, следующие условия:
— достаточное количество аминокислот
в клетке;
— запас энергии в клетке;
— активность ферментов и факторов
транскрипции РНК (РНК-полимераз и др.);
— высокий уровень анаболических
гормонов в крови (тестостерона и
соматотропина);
— наличие в клетке белков-рецепторов
тестостерона.
Теперь осталось лишь ответить на вопрос:
каким именно образом тренировка влияет на
синтез белка?
Я вынужден разочаровать читателя: детально
объяснить механизм этого влияния при
сегодняшнем уровне развития науки
невозможно. Если о том, как происходит
регуляция синтеза белка в простейших
одноклеточных организмах, в каждой клетке
которых идёт строительство всех белков,
закодированных в ДНК, учёные имеют
достаточно полное представление, то
регуляция синтеза белка в многоклеточных
организмах, когда теоретически каждая
клетка может синтезировать все возможные
белки, закодированные в ДНК, но
синтезирует лишь тот набор белков, который
присущ данному типу клеток, пока, увы,
остаётся загадкой. Да, гормоно-рецепторный
комплекс разблокирует тот участок ДНК,
где закодирован определённый белок, но
каким именно путём гормон узнаёт, какой
ген в данный момент необходим клетке
— миозин быстрого волокна или
миозин медленного волокна, а может быть,
миоглобин? Учёным ещё предстоит пройти
долгий путь, прежде чем раскроются все
тайны синтеза белка. А как же быть до тех
пор?
Сегодня существует несколько гипотез,
пытающихся объяснить влияние тренировки на
синтез белка в мышце. Но все эти гипотезы
можно объединить в два конкурирующих
направления: первое — теория накопления;
второе — теория разрушения.
Суть теории накопления состоит в том, что
во время мышечной деятельности в клетке
вырабатываются некие факторы-регуляторы,
оказывающие влияние на процессы считывания
информации с ДНК. Одни учёные относят к
этим факторам повышение кислотности среды
в результате мышечной деятельности,
влияющее на спирилизацию ДНК. Другие же
исследователи относят к
факторам-регуляторам свободный креатин:
при интенсивной деятельности креатинфосфат,
содержащийся в клетке, в целях восполнения
энергии передаёт фосфатную группу на АДФ,
превращаясь в креатин, и именно креатин, по
мнению упомянутых исследователей, оказывает
регулирующее воздействие на ДНК.
Я нисколько не сомневаюсь в том, что
подобные процессы и впрямь должны иметь
место в регуляции интенсивности белкового
обмена, — ведь, как известно, в случае
обездвижения мышцы интенсивность синтеза
белка в её клетках снижается, то есть
движение уже само по себе есть
фактор-регулятор белкового синтеза.
Впрочем, двигательная активность мышц является,
скорее, необходимым фактором, обеспечивающим
нормальный метаболизм в мышечной ткани, но
не достаточным условием для обеспечения
заметной гипертрофии мышц. Поскольку своё
регулирующее воздействие такие
факторы, как повышение уровня креатина или
кислотности среды, в наибольшей степени
оказывают во время работы мышц. В то время как
синтез белка идёт в основном после прекращения
нагрузки во время отдыха, когда концентрация
факторов-регуляторов уже
возвращается к уровню, характерному для
состояния покоя.
Куда более полное, на мой взгляд,
представление о механизмах регуляции синтеза
белков способна дать теория разрушения, суть
которой заключается в следующем.
Как я уже писал выше, организм — это
саморегулируемая система, настроенная
миллионами лет эволюции на поддержание
постоянства внутренней среды. Разрушение
внутренних структур организма автоматически
запускает процессы, направленные на
восстановление утраченного равновесия. Так,
разрушение белковых структур клетки в
здоровом организме активизирует
восстановительные процессы синтеза белка,
для протекания которых сразу же создаются
все необходимые условия. То, что активность
синтеза белка в повреждённой ткани в
несколько раз выше, чем в нормальных
условиях — это факт. Мало того,
интенсивные анаболические процессы не могут
затихнуть сразу по завершении
восстановления повреждённых структур. То
есть процессы синтеза белка, равно как и все
прочие процессы в организме, имеют некоторую
инерцию, так что их результатом почти всегда
бывает избыточный анаболизм, приводящий к
превышению уровня белка в клетке над
исходным. Другими словами, здесь тоже
происходит суперкомпенсация, уже
упоминавшаяся мной при рассказе о
восстановлении энергетических ресурсов. В
этом нет ничего особенного: восстановление
белковых структур клетки подчиняется общим
законам адаптации.
Обычно роль тренировки в гипертрофии мышц
исследователи сводят лишь к интенсификации
процессов синтеза РНК в ядрах клеток. Между
тем общий объём мышцы зависит от количества в
ней мышечных клеток и от количества ядер в этих
клетках. Согласно утвердившемуся в среде
спортивных физиологов представлению, число
мышечных клеток задаётся генетически и не
меняется в ходе тренировок, — об этом
свидетельствуют большинство экспериментов,
проводившихся в данном направлении (Б.С.Шекман).
Впрочем, имеется и ряд таких экспериментальных
данных, которые заставляют усомниться в
приведённом постулате (об этом чуть позже).
Объясняется неизменность количества клеток в
мышце тем, что ядра мышечной клетки
утрачивают способность к делению (равно как
и сама клетка) ещё на этапе эмбрионального
развития. Однако, как показывает ряд
экспериментов (M.Cabric и N.T.James), в ходе
тренировок количество ядер в мышечных
клетках всё-таки увеличивается.
Но как такое может происходить — ведь
разве ядра мышечных клеток не теряют навсегда
способность к делению? Откуда же тогда
появляются эти новые ядра?
Ответ на данный вопрос содержится в работах
учёных, занимавшихся проблемами регенерации
травмированной ткани. Как оказалось, на этапе
эмбрионального развития далеко не все клетки
эмбриона, из которых развивается затем
мышечная ткань, сливаются в мышечные волокна
и утрачивают способность к делению. Какая-то
часть из них (около 10%) остаётся в оболочке
волокон в виде клеток-сателлитов.
Клетки-сателлиты сохраняют способность к
делению на протяжении всей жизни организма и
являются резервом восстановления мышечной
ткани. А значит, только клетки-сателлиты
способны быть источником новых ядер в волокне.
Как показывают эксперименты (А.В.Володина;
Р.П.Женевская; А.А.Климов и Р.К.Данилов;
Э.Г.Улумбеков), повреждение мышечных волокон
приводит к активации клеток-сателлитов,
которые, освободившись из оболочки, вступают
в цикл деления, а затем сливаются воедино,
восстанавливая повреждённые волокна.
Логично предположить, что к активации
клеток-сателлитов после тренировки приводят
процессы, аналогичные травмам волокон.
Многие атлеты знают на собственном опыте,
что интенсивная тренировка, особенно после
продолжительного перерыва, отзывается болью
в последующие несколько дней отдыха. Боль
явно свидетельствует о разрушениях
внутренней структуры мышц. Тщательное
микроскопирование мышц показывает, что в
результате тренировок в ряде мышечных
волокон нарушается упорядоченное
расположение миофибрилл, распадаются
митохондрии, а в крови повышается уровень
лейкоцитов, как при травмах или при
инфекционном воспалении (В.И.Морозов;
М.Д.Штерлинг с соавторами). Разрушение
внутренней структуры мышечного волокна во
время тренировки (или, иными словами,
микротравма) приводит к появлению в волокне
обрывков белковых молекул, что сразу же
активизирует лизосомы, которые
"переваривают" с помощью содержащихся в них
ферментов подлежащие уничтожению белковые
структуры.
Если лизосомы не справляются с объёмом
повреждений, то примерно через сутки
наблюдается пик активности уже более
мощных "чистильщиков" — фагоцитов.
Фагоциты — это клетки, живущие в
межклеточном веществе и крови. Основная их
задача — уничтожение повреждённых
тканей и чужеродных микроорганизмов. Именно
продукты жизнедеятельности фагоцитов и
вызывают воспалительные процессы и боль в
мышцах примерно через сутки после
тренировки. По-видимому, как раз благодаря
деятельности лизосом и фагоцитов
повреждается оболочка мышечного волокна, и
из неё высвобождаются клетки-сателлиты.
Освободившись, клетки-сателлиты начинают
цикл деления и сливаются с повреждённым
волокном, увеличивая в нём количество ядер
и тем самым повышая его потенциал в плане
синтеза белка.
Учитывая все вышеизложенные обстоятельства,
я не стал бы полностью исключать возможность
высвобождения клеток-сателлитов в
межклеточное пространство и слияния их в
новые волокна (кстати, такое слияние
достоверно наблюдалось при обширных
повреждениях мышечной ткани — правда,
новые волокна в этом случае образовывались
только взамен утраченных, что, естественно,
не приводило к общему увеличению числа
волокон в мышце).
На мой взгляд, если повреждения мышечного
волокна не столь обширны, чтобы привести
его к гибели, а клетки-сателлиты тем не
менее всё же пошли по пути слияния в новое
волокно, то должна иметь место гиперплазия,
увеличение числа клеток. Тем более что
есть ряд экспериментов, выбивающихся из
общепринятых представлений о невозможности
гиперплазии. Так, W.Goneya удалось на 19-20%
увеличить количество мышечных волокон в
лапах кошек, которых он заставлял
тренироваться с прогрессирующей нагрузкой.
Мало этого, S.Yamada, N.Buffinger,
J.Dimario & R.Strohman (1989) и
L.Larson & P.A.Tesch (1986) сравнили
пробы мышечной ткани элитных бодибилдеров и
контрольной группы людей, обладавших
обычной мускулатурой, и этот анализ
показал, что поперечное сечение мышечных
клеток у элитных бодибилдеров было лишь
незначительно больше, чем у представителей
контрольной группы — в то время как
поперечное сечение самих мышц у
представителей обеих групп различалось
весьма существенно. То есть бодибилдеры
обладали заметно бОльшим количеством
волокон по сравнению с контрольной группой
— и это может быть объяснено либо
гиперплазией волокон, либо же тем, что
элитные бодибилдеры все как один имели
прямо с рождения аномально большое число
аномально тонких волокон (поскольку до
тренировок мускулатура у всех элитных
бодибилдеров была самых обычных объёмов).
Второе объяснение выглядит, безусловно,
не очень правдоподобным, поскольку требует
слишком уж многих и редких совпадений.
Впрочем, я не стану зацикливаться на
вопросе с гиперплазией и, поскольку
возможность последней у человека считается
недоказанной, буду исходить из того, что
рост мышц происходит исключительно по
причине гипертрофии уже существующих
волокон. Но ведь и одной из причин
гипертрофии мышечных волокон тоже является
увеличение в них числа клеточных ядер —
что по оказываемому эффекту сравнимо с
гиперплазией.
Вернусь к рассмотрению процессов,
происходящих в мышце во время восстановления
после тренировки. По завершении этапа
саморазрушения повреждённых тренировкой
структур начинается этап компенсации —
восстановления внутренней структуры волокон.
Конечно, данный этап не всегда заканчивается
суперкомпенсацией. При слишком обширных
повреждениях или отсутствии условий для
восстановления результат может быть
диаметрально противоположным.
Против теории разрушения чаще всего приводят
следующее возражение: "Если причиной роста
являются микротравмы, то почему же мышца не
растёт, когда её бьют палками?" Ответ на
данный вопрос можно найти в докторской
диссертации А.В.Володиной. Целью этой работы
являлось изучение процессов, препятствующих
реализации регенерационного потенциала,
заложенного в мышечном волокне. Эксперименты
показали, что в условиях обширного
повреждения волокон, сопровождающегося
ишемией (нарушением кровоснабжения)
повреждённых тканей, вызывающей дефицит в
снабжении волокна кислородом и питательными
веществами, одна часть клеток-сателлитов
гибнет и поглощается фагоцитами, а другая
часть идёт по пути превращения не в мышечные
клетки, а в фибробласты (клетки,
производящие коллаген). В итоге место
повреждения затягивается соединительной
тканью, а количество волокон в мышечной ткани
снижается по причине их частичной гибели от
повреждений.
При микротравмах же волокна (то есть при
повреждении одной лишь внутренней структуры
мышечной клетки без нарушения её целостности),
в отличие от травмы целой мышцы, снабжение
волокна кислородом, а также его иннервация не
страдают — следовательно, в этом случае
нет факторов, приводящих к гибели
клетки-сателлиты и целые волокна.
В общем, если объём микротравм, полученных в
ходе тренировки, не слишком велик для срыва
восстановительных процессов, но достаточен
для активации клеток-сателлитов, то в
подвергшихся тренировочной нагрузке волокнах
увеличивается количество клеточных ядер.
Восстановление энергетических ресурсов после
тренировки приводит к суперкомпенсации
энергетических структур, а лизис разрушенных
тренировкой белков увеличивает содержание
свободных аминокислот непосредственно в
волокнах, что в совокупности создаёт
благоприятные условия для интенсификации
процессов синтеза белка. При условиях
достаточного по времени отдыха, отсутствия
новых стрессовых нагрузок, адекватного
снабжения мышечных клеток энергией и
пластическими ресурсами (аминокислотами)
интенсивные процессы восстановления приводят
к накоплению в волокнах белковых структур
сверх того уровня, который имел место до
тренировки — то есть происходит
гипертрофия мышц.
Надо отметить, что последовательность
протекания фаз общей неспецифической
адаптационной реакции (синдрома стресса)
такова, что обеспечивает поддержку
описанных выше регенерационных процессов
на системном уровне. Первая катаболическая
фаза стресс-реакции сопровождается
выбросом кортикостероидов, что приводит
к мобилизации энергетических ресурсов
организма и обеспечивает выработку
ферментов лизосом и фагоцитов, расщепляющих
белок (кортикостероиды являются теми
гормонами, которые активируют у клеток
гены протеолитических ферментов), что
способствует скорейшему очищению волокон
от повреждённых структур. В последующей
фазе стресс-реакции синтез кортикостероидов
сменяется синтезом анаболических гормонов,
что обеспечивает на системном уровне
компенсаторный анаболизм.
Что такое микротравма?
Остаётся открытым следующий вопрос: что
вызывает разрушение внутренней структуры
волокна и является, тем самым, стрессом
для мышцы? Прежде чем ответить на этот
вопрос, придётся вспомнить механизм
сокращения мышц, описанный в I части
данного текста.
Учёный и пауэрлифтёр Фредерик Хэтфилд,
считающий роль микротравм в тренировочном
процессе, скорее, отрицательной из-за
необходимости длительного восстановления,
полагает, что причиной микротравм
является повреждение миофибрильных нитей
во время негативных повторений. Вот как
Хэтфилд описал механизм таких повреждений:
"Поскольку количество перекрёстных
мостиков, старающихся сократить мышцу,
недостаточно, они буквально "продираются"
сквозь мостики соединений нити, стараясь
вызвать концентрическое сокращение.
Однако сцепиться как следует им не
удаётся, они срываются и повреждаются.
Эти действия, очень напоминающие
протаскивание щетины одной зубной щётки
через другую, сопровождаются сильным
трением, и мышечные нити разрушаются."
Не правда ли, образно? Но, на мой взгляд,
несколько сумбурно: так пишут, когда
хотят объяснить то, что до конца не
понимают сами. К сожалению, Ф.Хэтфилд в
своём объяснении ошибся дважды.
Во-первых, микротравмы возникают не
только при негативных повторениях, но и
при позитивном движении, что легко можно
наблюдать на практике, делая, например,
становые тяги с полностью исключённой
негативной фазой движения.
Во-вторых, использовать термин "трение"
для описания взаимодействия молекул
— некорректно. Понятие силы трения
введено в физике для описания на
макроуровне поверхностного взаимодействия
тел специально для того, чтобы
абстрагироваться от истинной природы
"трения": электромагнитного взаимодействия
молекул поверхностного слоя.
На самом деле механизм повреждения
миофибрильных нитей вовсе не таков, каким
представил его Хэтфилд; мне удалось
смоделировать этот механизм в результате
более детального изучения биохимических
процессов, происходящих при сокращении
мышечной клетки.
Для понимания того, как происходит
повреждение миофибрильной нити, читателю
придётся ещё раз, — причём теперь уже
более подробно — ознакомиться с
фазами движения миозинового
мостика (рис. 8).
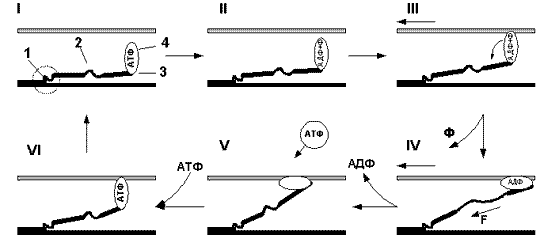
Рис. 8 (1 и 3 — шарнирные участки, 2 — эластичный компонент, 4 —
глобулярная головка миозина) |
Молекула миозина состоит из лёгких и
тяжёлых цепей меромиозина. Тяжёлая цепь
меромиозина содержит в себе две
глобулярные головки (4) (на рисунках для
простоты изображена только одна головка),
и обе эти головки через шарнирный
участок (3) связаны с эластичным
компонентом (2). Лёгкая цепь меромиозина
соединена с тяжёлой цепью шарнирным
участком (1).
В первой фазе, ещё до сцепления с актином,
головка миозинового мостика несёт в
себе АТФ. Во второй фазе под действием
фермента АТФазы, локализованного в самой
головке миозина, АТФ гидролизуется и
расщепляется на АДФ и неорганический
фосфат, причём происходит это на не
связанном с актином миозине. После чего,
благодаря повороту в шарнирном участке 1,
миозиновая головка соединяется с актином
— что является третьей фазой
процесса. После соединения с актином
головка миозина проворачивается в
шарнирном участке 3, благодаря чему
происходит натяжение эластичного
компонента, и мостик генерирует усилие.
Для поворота головки и совершения
рабочего хода мостика используется
энергия, освобождённая при диссоциации
продуктов гидролиза АТФ. Основная доля
этой энергии выделяется при высвобождении
неорганического фосфата (переход из
третьей фазы в четвёртую), а меньшая часть
— при высвобождении АДФ (переход из
четвёртой фазы в пятую). В пятой фазе
возникает сцепленное или "ригорное"
состояние мостика, то есть мостик уже не
генерирует движущую силу, но остаётся
сцепленным с актином, и вывести его из
этого состояния может только молекула АТФ.
Поглощая АТФ, головка миозина переходит в
шестую фазу, после чего отцепляется от
актина, возвращаясь в исходное состояние
(первую фазу).
Разбираясь с фазами движения миозинового
мостика, я сразу обратил внимание на тот
факт, что для отцепления мостика от
актина требуется молекула АТФ. При
скольжении нитей миозина вдоль актина
под действием сил тянущих мостиков (в
случае позитивного движения) или под
действием внешней силы (в случае негативного
движения) сцепленные мостики растягиваются и
мешают движению — этим, как я уже
писал, и объясняется различие в силе,
развиваемой волокном при удлинении и при
сокращении, а также при сокращениях с
разной скоростью. Понятно, что когда АТФ
в мышце содержится в достаточном
количестве, все мостики успевают
отцепиться вовремя — но что должно
происходить при очень сильном снижении
концентрации АТФ в мышце? Несомненно, тут
должно происходить следующее: молекула АТФ
вполне может не успеть отцепить головку
миозина до того, как растяжение мостика
превысит предел его прочности — и
сцепленный мостик тогда, естественно,
разорвётся (рис. 9). Разумеется, место
разрыва мостика изображено мной
достаточно условно, я не могу точно
указать, где находится самое слабое звено
в цепи.
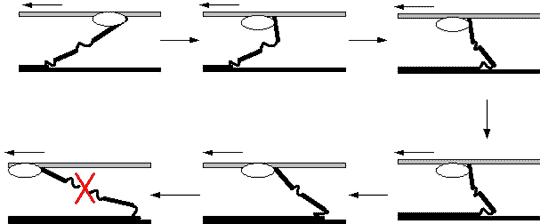
Рис. 9 |
Все, наверное, слышали о состоянии
трупного окоченения мышц. Наступает оно
именно потому, что в мёртвом организме
запасы АТФ не восполняются, и мостики
миозина оказываются накрепко сцепленными
с актином. Что произойдёт с мышцей
трупа, если её насильно растянуть?
Понятно, что — разрыв. Так вот нечто
подобное должно происходить и с
отдельными волокнами живой мышцы при
растяжении, если в них произойдёт резкое
снижение уровня АТФ.
Интересно, что при недостатке АТФ
наиболее разрушительным воздействием на
мышцы должна обладать негативная фаза
движения. Как я уже писал, мостик
генерирует силу благодаря натяжению
эластичного компонента после проворота
головки миозина ("гребка"). В случае
позитивного движения скольжение нитей
после "гребка" поначалу приводит к
ослаблению натяжения эластичного
компонента, и только дальнейшее смещение
нитей под действием других мостиков может
привести к последующему растяжению
данного "ригорного" мостика. В случае же
негативного движения скольжение нитей
под действием внешней силы происходит в
сторону, противоположную направлению силы
упругости эластичного компонента мостика,
поэтому относительное смещение нитей
после "гребка" сразу же приводит к ещё
большему натяжению эластичного компонента
"ригорного" мостика (рис. 10). Таким
образом, вероятность вынужденного разрыва
актино-миозинового комплекса в
условиях низкого уровня АТФ значительно выше в
негативной фазе движения.
Проведённый анализ показывает, что
причиной микротравм мышц при тренировках с
большой вероятностью может быть именно
снижение уровня АТФ в ряде активно
работающих волокон. Тогда становится
понятным, почему большинство специалистов
возлагают вину за повреждение мышц именно
на негативную фазу движения.
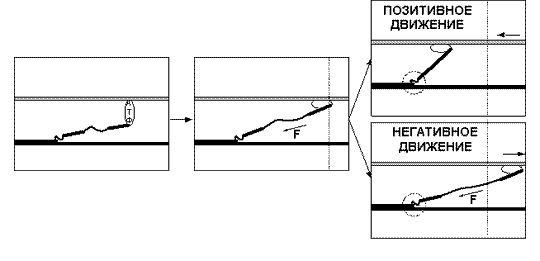
Рис. 10 |
Некоторые специалисты (например, Мак-Комас)
видят причину повреждения мышц при
негативном движении в микроразрывах
оболочки волокон при её растяжении. На
первый взгляд, волокно, а следовательно, и
его оболочка, могут быть растянуты лишь
настолько, насколько позволяют суставы и
связки (естественный ход мышцы). Никакого
растяжения оболочки волокна, приводящего
к микротравмам, при движении без нагрузки
не возникает. Почему же негативное
движение под нагрузкой должно приводить к
растяжению оболочки мышечных волокон?
Мак-Комас привёл в своей книге мнение, что
в условиях негативного движения отдельные
саркомеры могут проявлять нестабильность
и растягиваться быстрее остальных, в
результате принимая длину большую, чем они
имеют при полном растяжении волокна в
нормальных условиях.
Рис. 11 |
Если проведённый мной анализ поведения
актино-миозинового комплекса в условиях
низкого уровня АТФ является недостаточным
для объяснения разрушительного воздействия
негативных повторений, если нестабильность
и сверхрастяжение саркомеров действительно
наблюдались на практике (из текста
Мак-Комаса неясно, зафиксировано в
эксперименте именно сверхрастяжение
саркомеров или же оно было привлечено
просто для теоретического объяснения
наблюдаемого факта повреждения волокон),
то что же тогда делает растяжение
саркомеров нестабильным?
Примечательно, что микротравмы волокон,
а соответственно, и возможная
нестабильность саркомеров проявляются не
при любом негативном движении, а лишь
при достаточно интенсивном и длительном.
Смею предположить, что причина
нестабильности саркомеров может крыться
также в критическом снижении уровня АТФ
в мышечных волокнах, приводящем к
ригидности (или некоторому замедлению
растяжения) части саркомеров. Растяжение
волокна до его исходных размеров в этих
условиях может привести к
сверхрастяжению остальных (неригидных)
саркомеров (рис. 11).
Итак, то, что при движении,
сопровождающемся недостатком АТФ,
нарушается нормальное сокращение-растяжение
актино-миозиновых нитей и возникают
внутренние напряжения, приводящие к
микротравмам волокна, — неоспоримо.
Но дабы быть уверенными в том, что
микротравмы мышц, возникающие в ходе
тренировок, действительно имеют именно
ту природу, которую я предложил,
необходимо понять, возможно ли в ходе
тренировки соответствующее снижения
уровня АТФ в мышечных волокнах, а если
возможно, то при каких условиях.
Как мне показалось поначалу, приведённые
соображения должны служить исчерпывающим
объяснением действенности системы Менцера:
работа до "отказа" приводит к истощению
запасов АТФ и, соответственно, к разрывам
в актино-миозиновом комплексе, что
становится стрессом для мышцы и вызывает
адаптационную реакцию.
Однако на самом деле данная "отказная"
схема генезиса микротравмы весьма далека
от реальности: как выяснилось, в
большинстве случаев "отказ" наступает
вовсе не из-за исчерпания
запасов АТФ
— эксперименты показывают, что
уровень АТФ может оставаться
достаточно высоким даже в уставшей мышце.
Усомниться в правильности "отказной"
схемы меня заставил ещё и тот факт, что
появление боли в мышцах — этого
вестника микротравм — не имеет
однозначной связи с работой до "отказа".
Ведь, с одной стороны, на начальном
этапе тренинга боль в мышцах возникает
почти при любой более-менее интенсивной
работе, то есть независимо от того, был
"отказ" или нет. А с другой стороны,
регулярные тренировки приводят к тому,
что работа даже сверх "отказа" (читинг,
"стриптиз" и др.) не вызывает уже
никакой посттренировочной боли —
боль возвращается только после
продолжительного перерыва в тренировках.
Все эти противоречащие первоначально
найденному "решению" факты и вынудили
меня заняться изучением феномена "отказа".
Что такое "отказ" мышцы
"Отказ" — понятие довольно условное.
Это не какое-то особое состояние мышцы,
а просто её неспособность развить силу,
необходимую для преодоления внешней
нагрузки.
Факторы утомления, приводящие к снижению
силы мышечного сокращения, принято
разделять на центральные и периферические.
Центральное утомление развивается в
нейронах коры головного мозга,
генерирующих импульс к сокращению, а
также в нисходящих путях и двигательных
мотонейронах спинного мозга, и выражается
в снижении количества активных
двигательных мотонейронов и в уменьшении
частоты их импульсации. Развитие утомления
в двигательных центрах коры головного
мозга принято называть "охранительным
торможением".
Периферическое утомление проявляется в
ухудшении передачи сигнала с аксона на
мышечное волокно (утомление в
нервно-мышечном синапсе) и в снижении
силы сокращения самих волокон, даже в
условиях их нормальной активации.
Причиной нарушения передачи сигнала в
нервно-мышечном синапсе является
накопление в мышце ряда продуктов
метаболизма при её двигательной
активности (Д.Ю.Романовский). Но даже
если сигнал от мотонейрона достигает
поверхности мышечных волокон и вызывает
возбуждение мембраны волокна,
полноценное сокращение волокна ещё не
гарантировано, так как накопление
метаболитов при высокочастотной
стимуляции волокна приводит к нарушению
распространения сигнала от поверхности
волокна к терминальным цистернам
внутренних миофибрилл, в результате чего
в глубоко лежащих миофибриллах кальций
перестаёт выбрасываться в саркоплазму, и
миозиновые мостики не имеют возможности
сцепляться с актином и генерировать силу
— то есть наблюдается так называемое
нарушение сочетания
"возбуждение-сокращение" (Дж.Мак-Комас).
Мак-Комас считает нарушение сочетания
"возбуждение-сокращение" ведущим фактором
периферического утомления при интенсивных
сокращениях.
Но, кроме факторов утомления, связанных с
нарушениями процессов генерации и
распространения нервного импульса,
существуют и иные причины, приводящие к
снижению силы, развиваемой мышечными
волокнами — даже в условиях их
полной активации. Как можно видеть из
модели скользящих нитей, сила отдельного
волокна зависит от количества
одновременно тянущих миозиновых мостиков.
Биохимических причин снижения силы,
развиваемой мостиком в генерирующих силу
фазах (III, IV) не существует (во всяком
случае, пока об этом ничего не известно),
поэтому сила мышечного волокна (при его
достаточном возбуждении) может снизиться
только по следующим причинам:
1) из-за увеличения времени пребывания
мостиков в ригорном (сцепленном, не
генерирующем силу) состоянии (фаза V, VI);
2) из-за увеличения времени пребывания
мостиков в расцепленном состоянии (фаза I, II).
Время нахождения мостика в ригорном
состоянии зависит от того, насколько
быстро АТФ вступает в контакт с головкой
миозина. Время это равно частному от
деления расстояния (h) от мостика до
ближайшей молекулы АТФ, движущейся в
направлении мостика (ведь, как известно,
молекулы постоянно находятся в
хаотическом движении), на скорость движения
этой молекулы (v). А среднее время (t)
пребывания мостиков в ригорном состоянии
равно, понятно, средневзвешенному
расстоянию между молекулами АТФ и мостиком,
делённому на средневзвешенную скорость
движения молекул. То есть
t = h/v
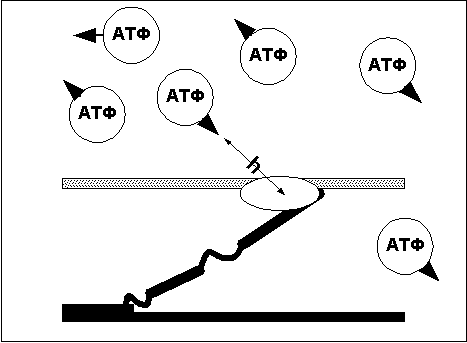
Рис. 12 |
Понятно также, что чем выше концентрация АТФ
в мышце, тем меньше среднее расстояние
между молекулами АТФ и головками миозина
и, соответственно, тем меньше время
нахождения мостиков в ригорном состоянии.
А это значит, что сила волокна зависит
от концентрации АТФ. То есть при прочих
равных условиях волокно с высокой
концентрацией АТФ будет сильнее такого
же точно волокна, но с меньшей
концентрацией АТФ. Как известно,
воспроизводство АТФ за счёт креатинфосфата
происходит почти мгновенно, поэтому можно
с уверенностью утверждать, что
концентрация АТФ при работе мышцы зависит
от концентрации креатинфосфата.
Итак: чем выше концентрация
энергетических фосфатов (АТФ и
креатинфосфата) в мышечном волокне, тем
выше сила, развиваемая этим волокном.
Концентрация энергетических фосфатов
оказывает второе по значимости влияние
на силу мышечного волокна (при условии
его полной активации) после такого
фактора, как количество и поперечное
сечение миофибрилл. Это положение имеет
прямое практическое подтверждение:
атлеты, повышающие уровень содержания
креатинфосфата в мышцах за счёт приёма
креатина в качестве пищевой добавки,
отмечают заметную прибавку в силе.
Зависимость силы волокна от концентрации
энергетических фосфатов не является
линейной. При высоких концентрациях, когда
время ожидания контакта АТФ с
головкой миозина становится меньшим или
сопоставимым со временем, необходимым для
протекания химической реакции, приводящей к
отделению головки миозина от актина,
повышение концентрации АТФ и
креатинфосфата уже мало влияет на скорость
переключения мостиков и, соответственно,
на силу сокращения волокна. Зато низкие
концентрации АТФ и
креатинфосфата существенно снижают силу
сокращения волокна, что может служить одной
из причин "отказа".
Теперь подошла очередь рассмотреть второй
фактор, уменьшающий силу сокращения
волокна: увеличение времени пребывания
мостиков в разомкнутом состоянии
(в фазах I и II). Последнее чаще всего
происходит в результате снижения скорости
гидролиза АТФ и расщепления её
на АДФ и
неорганический фосфат. В свою очередь,
снижение скорости гидролиза является, как
правило, следствием понижения
активности АТФазы миозина (фермента,
катализирующего реакцию гидролиза АТФ).
Что же может понижать активность АТФазы?
Расщепление АТФ и его воспроизводство
за счёт креатинфосфата вызывает
накопление в мышце ортофосфорной
кислоты, гликолиз ведёт к повышению
концентрации в саркоплазме молочной
кислоты, окисление же, в свою очередь,
"отравляет" мышцы угольной кислотой.
Активность АТФазы как раз очень сильно
зависит от кислотности среды. Как
показывают эксперименты, максимум
АТФазной активности достигается в среде,
близкой к нейтральной (рН = 7). При
смещении pH среды мышцы в кислую
сторону активность АТФазы снижается, а при
падении рН мышцы до пяти
АТФазная активность близится к нулю (П.Г.Богач
с соавторами; Б.Ф.Поглазов). Таким образом, в
результате накопления кислых продуктов
метаболизма АТФаза миозина постепенно
снижает скорость гидролиза АТФ, и
миозиновые мостики теряют способность
сцепляться с актином — то есть мышца
уменьшает силу сокращения, несмотря даже
на поступающий от мотонейрона сигнал.
Таким образом, "отказ" мышцы является
результатом воздействия сразу нескольких
факторов утомления, доля каждого из
которых в конечном счёте определяется
интенсивностью и длительностью работы мышцы.
При этом снижение концентрации АТФ в мышце,
хотя и наблюдается в небольшом диапазоне
с 5 ммоль/кг до 3 ммоль/кг (Н.И.Волков),
не является доминирующим фактором утомления,
и "отказ" мышцы может возникать даже при
высоком уровне АТФ. Больше того, все
механизмы мышечного утомления направлены на
прекращение мышечной активности именно до
истощения запасов энергии — для
предотвращения необратимых последствий для
мышц и всего организма.
Но то, что при утомлении не фиксируется
кардинальное снижение концентрации АТФ в
мышце, ни в коей мере не может быть
аргументом в пользу того, что снижение
концентрации АТФ невозможно ни при каких
условиях. Во-первых, в тех экспериментах,
в которых проводились измерения
концентрации АТФ, как правило, фиксировалось
среднее значение уровня АТФ по мышце —
а это всё равно что средняя температура
больных по палате. При определённом
количестве АТФ в пробе мышечной ткани
концентрация АТФ в отдельных волокнах, а уж
тем более в отдельных участках волокон
может сильно отличаться от среднемышечного
показателя. Во-вторых,
биопсия (проба) утомлённой мышечной ткани
совершалась уже после прекращения
движения, а поскольку скорость
метаболических процессов крайне высока,
то измерения показывали уровень АТФ не в
сокращающейся мышце, а в мышце через
некоторое время (пусть и весьма короткое
по человеческим меркам) после прекращения
работы. В-третьих (и это главное),
практически все биохимические эксперименты
по измерению уровня АТФ традиционно
проводились на мышцах, утомлённых на
велотренажёре, беговой дорожке и
аналогичных лабораторных установках, между
тем как после нагрузки такого рода
микротравмы мышц появляются весьма редко
— а это значит, что существенное
снижение уровня АТФ в подобных упражнениях
вряд ли происходит.
Таким образом, относительное постоянство
средней концентрации АТФ в мышце,
обнаруживаемое в экспериментах, вовсе не
исключает возможность локального временного
снижения уровня АТФ в отдельных саркомерах.
Для ответа на вопрос, при каких условиях
проявление такой нестабильности в
концентрации АТФ наиболее вероятно, следует
обратиться к рассмотрению
последовательности активации метаболических
процессов в мышцах.
Ещё раз об энергетике
Резкий переход к интенсивной мышечной
деятельности из состояния покоя требует
столь же резкого увеличения скорости
производства энергии. Для достижения
максимальной мощности основных источников
воспроизводства энергии (гликолиза в
быстрых волокнах и окисления в медленных)
требуется определённое время. Так,
скорость воспроизводства АТФ за счёт
гликолиза достигает максимума только
через 20-30 секунд после начала
интенсивной работы. Хотя, по некоторым
данным, может происходить и более быстрая
активация гликолиза (до 5 секунд), а
столь медленное его разворачивание в
обычных условиях связано с тем, что
гликолиз не получает достаточных стимулов
для развёртывания, пока сохраняется
высокий уровень креатинфосфата в мышцах.
Для достижения же максимальной скорости
окислительного процесса требуется ещё
больше времени, и связано это в основном
с необходимостью оптимизации
процессов доставки кислорода. Таким
образом, скорость окисления становится
максимальной лишь через 1-2 минуты
после начала работы мышц. Между тем
мышца развивает максимальную мощность
с первых же секунд после поступления
сигнала к сокращению. Причём мощность
оного сокращения такова, что гликолиз
вкупе с окислением (даже при условии
максимальной активации данных источников)
не в состоянии обеспечить скорость
воспроизводства АТФ, необходимую для
поддержания этой наивысшей мощности.
Приведение в соответствие скоростей
расходования и воспроизводства АТФ во время
работы мышцы идёт сразу по двум
направлениям. Во-первых, постепенная
активизация гликолиза и окисления
увеличивает количество АТФ, синтезируемого
в единицу времени за счёт этих источников.
Во-вторых, накопление продуктов
метаболизма в результате интенсивной
мышечной деятельности приводит к развитию
мышечного утомления и снижению силы
сокращения и, соответственно, скорости
расходования энергии. Благодаря этим двум
процессам, скорости расходования и
воспроизводства АТФ выравниваются, и в
дальнейшем работа идёт уже с постепенно
снижающейся мощностью, но зато с
сохранением равновесия между количеством
потребляемого и синтезируемого АТФ.
Таким образом обеспечивается стабильность
уровня АТФ в мышце и "отказ" наступает не
вследствие истощения запасов АТФ,
а из-за снижения сократительной
способности мышц в результате развития описанных
выше процессов утомления.
До выравнивания скоростей расходования и
воспроизводства энергии дефицит АТФ
восполняется за счёт имеющегося в мышце
креатинфосфата. То есть креатинфосфат
играет роль буфера, сглаживающего
разницу в скоростях воспроизводства и
потребления АТФ при резко возрастающих
нагрузках и обеспечивающего постоянство
концентрации АТФ. В обычной жизни мы
редко используем мышцы на пределе их
энергетических возможностей, поэтому
они вполне успешно обходятся небольшим
запасом креатинфосфата и ферментов,
обеспечивающих протекание реакций
гликолиза и окисления. Но если дать
нагрузку, значительно превышающую
привычную, то запас креатинфосфата в
наиболее активно работающих волокнах
очень быстро иссякнет — задолго до
того момента, когда скорости потребления
и производства АТФ за счёт основных,
более ёмких источников придут в
равновесие.
Таким образом, при низком уровне
креатинфосфата, слишком интенсивном
расходовании энергии и неадекватной
скорости её воспроизводства и становится
возможным падение уровня АТФ ниже
критического (обеспечивающего
своевременное отцепление головок миозина
от актина) — причём не только в
отдельных саркомерах, но также и в целых
волокнах. В этих условиях продолжение
сокращения миофибрилл под действием
активных мостиков (при позитивном
движении) либо их, миофибрилл, растяжение
под действием внешней силы (при
негативном движение), как я показал чуть
выше, должно приводить к разрушению
отдельных цепей миозина и даже целых
волокон.
Итак, вот он, главный секрет
тренировочного стресса: микротравмы
мышечного волокна возникают при условии
критического понижения в нём запасов
креатинфосфата.
Именно этим и объясняется тот факт, что
эффект наиболее разрушительного
воздействия на быстрые волокна
достигается при интенсивной работе
длительностью от семи до тридцати секунд.
Если нагрузка позволяет поддерживать
сокращение мышц дольше тридцати секунд,
то расходование энергии в относительно
тренированной мышце, скорее всего, будет
недостаточно интенсивным для того, чтобы
концентрация АТФ и креатинфосфата упала
ниже критического уровня. В свою очередь,
когда нагрузка бывает велика настолько,
что работа может продлиться не дольше
нескольких секунд (одно-два повторения),
имеет место совсем иная картина. Скорость
расходования энергии в этом случае
оказывается очень высока, но "отказ",
вызываемый уже самым незначительным
уменьшением силы волокон, происходит ещё
до момента исчерпания запасов
креатинфосфата — и стрессовая ситуация
тем самым просто не успевает сложиться.
Понятно, что при максимально интенсивном
режиме работы мышц добиться микротравм в
окислительных (медленных) волокнах совершенно
невозможно — ведь скорость
расходования АТФ в медленных волокнах
значительно ниже, чем в быстрых, поэтому запасы
креатинфосфата истощаются слишком плавно. Получение
микротравм в медленных волокнах было бы,
пожалуй, практически нереальным, если для
активации окислительных процессов
требовалось бы столько же времени, сколько его
требуется для активизации гликолиза. Однако,
как я упоминал ранее, максимум выработки АТФ
за счёт окисления достигается через 1-2 минуты
работы, поэтому есть шанс добиться микротравм
даже и в медленных волокнах — нужно
только успеть получить дефицит креатинфосфата,
а затем и АТФ в результате интенсивной работы
в течение 1-2 минут до того, как
выработка АТФ в медленных
волокнах достигнет максимальных значений.
То, что окислительные волокна энергетически
более устойчивы и меньше подвержены
микротравмам, на мой взгляд, и является
объяснением того фиксируемого большинством
спортивных исследователей факта, что
гипертрофия мышц реализуется в основном
благодаря гипертрофии быстрых волокон.
Предложенная мною модель возникновения
микротравм неплохо согласуется ещё с одним
физиологически важным явлением, известным
каждому спортсмену, но так до сих пор и не
получившему сколько-нибудь приемлемого
объяснения: посттренировочная боль особенно
сильна после первых занятий и практически
полностью исчезает при регулярных
тренировках — но появляется вновь в
случае длительного перерыва в тренировочных
нагрузках. Последнее явление с позиции
приведённой выше модели объясняется очень
просто: реакцией организма на тренировку,
помимо усиления синтеза белка, является
также накопление в мышце креатинфосфата и
повышение концентрации и активности
ферментов гликолиза и окисления.
Соответственно, добиться исчерпания
возросших запасов креатинфосфата до
выравнивания скоростей расходования и
восстановления АТФ за счёт основных
источников энергии становится сначала всё
труднее, а затем уже и просто невозможно,
поскольку сокращение мышц становится
энергетически всё более и более устойчивым.
А вот и ещё один очевидный вывод:
накопление креатинфосфата и рост мощности
гликолиза и окисления в результате
тренировок, с одной стороны, повышают силу
мышц и способствуют росту их
работоспособности, но, с другой стороны,
препятствуют созданию стрессовых ситуаций
и снижают воздействие тренировки на мышцу,
замедляя тем самым дальнейшие
адаптационные реакции.
Явление постепенного "привыкания" мышц к
нагрузке уже давно известно специалистам
бодибилдинга под названием "тренировочное
плато". Но вот сами биохимические причины
процессов, приводящих к снижению
восприимчивости мышц к тренировке, были
неизвестны. Поэтому для выхода из
тренировочного плато специалисты чаще
всего советовали сменить режим работы,
заменить выполняемые упражнения, изменить
тренировочный сплит либо увеличить объём
нагрузки, чтобы как-то по-новому
воздействовать на мышцу и добиться от неё
реакции.
Между тем для того, чтобы достигнуть
снижения концентрации АТФ в условиях
накопления в мышце всё большого и
большего количества креатинфосфата и
роста её энергообменных возможностей,
необходимо повышать скорость расходования
энергии — для чего в распоряжении
атлета имеется не так уж и много
способов. Один из них — повышение
веса снаряда. Однако постоянное
увеличение веса снаряда с целью
интенсифицировать нагрузку приводит к
тому, что количество повторений в
упражнении опускается ниже четырёх,
что, как я отмечал выше, не может
оказать на мышцу необходимого воздействия.
Увеличение же объёма работы за счёт роста
количества упражнений и подходов не
всегда оказывается эффективным — ведь
если интенсивность расходования энергии
недостаточно высока для исчерпания
накопленного в мышце креатинфосфата до
выравнивания скоростей расходования и
синтеза АТФ и стрессовая ситуация не
наступает в первом же подходе, то
последующие подходы, скорость расходования
энергии в которых меньше, чем в первом
подходе, ввиду остаточного накопления
продуктов метаболизма тем более не дадут
нужного эффекта. (Напоминаю, что уровень
креатинфосфата в мышце восстанавливается
в течение нескольких минут, в то время
как молочная кислота, один из главных
метаболитов утомления, полностью
выводится из мышцы лишь через несколько
часов после тренировки.) Таким образом,
по мере приспособления мышц к нагрузкам
тренировка из стрессового фактора
превращается в обычную работу. То есть
теперь спортсмен может тренироваться хоть
каждый день, не испытывая особой
перетренированности, поскольку для
восстановления мышц ему оказывается
достаточным всего лишь одного-двух дней
отдыха — именно потому, что привычная
тренировка не вызывает разрушения волокон.
Есть ли, однако, смысл в подобного рода
тренировках? Безусловно, есть: ведь при
постепенном наращивании объёма работы
будет расти, например, объём саркоплазмы
волокон — за счёт накопления
энергетических веществ. Однако подобный
рост мышц далеко не беспределен. Без
увеличения количества и объёма миофибрилл,
а особенно количества клеточных ядер в
волокнах (кстати, не стоит исключать тут
и возможность увеличения количества самих
волокон), добиться значительной гипертрофии
мышц просто невозможно.
Как же быть, неужели это тупик?
Часть III. Критический анализ
тренировочных методик
Во второй части данного текста я показал, что
причиной роста функциональных возможностей мышц
и, в частности, причиной гипертрофии мышечной
ткани являются разнообразные адаптационные процессы,
происходящие в организме в ответ на вызванное
тренировкой изменение внутренней среды мышц, и
что именно ряд этих же адаптационных процессов
приводит к постепенному снижению амплитуды
возмущения внутренней среды в ответ на нагрузку
и, соответственно, к снижению интенсивности
дальнейших адаптационных процессов. Что в
конечном счёте и приводит к остановке
тренировочного прогресса — то есть приводит к
состоянию, называемому "тренировочное плато".
Как же избежать "привыкания" мышц к нагрузке,
как добиться постоянного прогресса в тренировках?
"Супертренинг" Менцера
Похоже, одно из решений проблемы в рамках
бодибилдинга найдено Майком Менцером, хотя
исходил он из несколько иных предпосылок.
Наблюдая за учениками, Менцер заметил, что
при принятой в бодибилдинге частоте тренировок
(2-3 тренировки в неделю
на одну группу мышц) первоначальный прогресс очень
быстро прекращается и наступает застой.
Полагая, что причина в "перетренированности"
— то есть в срыве восстановительных
способностей организма — Менцер стал
экспериментировать с уменьшением объёма
нагрузок и увеличением времени отдыха. Итогом
его экспериментов стал знаменитый "Супертренинг"
— тренировки одной группы мышц не чаще, чем
раз в одну-две недели в одном подходе.
При таких тренировках прогресс, по утверждению
Менцера, идёт непрерывно до полного исчерпания
потенциала атлета. С "классической" точки
зрения на тренировочный процесс, при столь
редких и малообъёмных тренировках вообще
невозможен сколько-нибудь заметный
прогресс.
Так в чём же секрет "Супертренинга"?
Эксперименты над лабораторными животными
(А.Ф.Краснова, 1960 г.) показали, что
после прекращения тренировок концентрация
креатинфосфата, гликогена, ферментов,
ускоряющих реакции энергопроизводства,
возвращается к дотренировочному уровню уже
через 1-3 недели отдыха, а вот
содержание сократительных белков в мышечном
волокне остаётся на достигнутом тренировкой уровне
ещё 30-40 дней после прекращения
тренировок. Таким образом, структурный след
адаптационных реакций, проявляющийся в виде
накопления сократительных белков мышц,
является достаточно стойким, а так называемая
"потеря формы" при перерыве в тренировках в
краткосрочной перспективе является следствием
снижения энергетического потенциала мышцы.
Соответственно, рекомендуемый Менцером отдых
между тренировками одной мышцы (1-2 недели)
не может привести к дезадаптации сократительных
структур мышц, чего как огня боятся
спортсмены. В то же время такой длительный
перерыв в тренировках не создаёт условий для
закрепления возникающих в ходе отдельных
тренировок адаптационных изменений в
энергетике мышц. Тем самым избирательно
делается акцент на самой важной для
бодибилдера адаптационной реакции — на
накоплении сократительных структур, и
искусственно сдерживаются другие
адаптационные процессы, приводящие к снижению
восприимчивости мышц к нагрузке. Итак,
оказывается, что столь длительный перерыв
между тренировками в "Супертренинге"
необходим не только для обеспечения
максимально полного восстановления мышц после
нагрузки, как полагал Менцер, но и для
предотвращения полной адаптации мышц, что
обеспечивает регулярное создание стрессовых
ситуаций. По степени воздействия на мышцу
каждая последующая тренировка в "Супертренинге"
Менцера практически равносильна первой.
В своей книге Менцер утверждает, что атлет,
несколько лет интенсивно тренировавшийся
4-5 раз в неделю, сможет получить эффект от
тренировок по его системе только после не
менее чем месячного отдыха. Столь
продолжительный перерыв в тренировках, по
моему мнению, как раз и необходим для
понижения "тренированности" мышц. Сам же
Менцер считал, что длительный отдых необходим
исключительно для восстановления организма, а
причина остановки в росте массы кроется в якобы
хронической "перетренированности" спортсменов,
тренирующихся по классическим методикам. Не
вникая в физиологию мышечной деятельности, но
опытным путём уловив некоторые основные
тенденции, Менцер сделал ещё ряд ошибочных
выводов.
Например, утверждение Менцера о том, что креатин
(как пищевая добавка) замечательно работает
в его системе, является, скорее всего,
вынужденным рекламным заявлением. При столь
редких тренировках, как в "Супертренинге",
ни о каком накоплении креатинфосфата в мышцах
не может быть и речи, поскольку его уровень
определяется прежде всего регулярностью
тренировок, а не количеством креатина,
потребляемого с пищей. Более того, как я
показал во второй части, эффект "Супертренинга"
как раз и заключается в поддержании
креатинфосфата в мышцах на относительно низком
уровне.
Рекомендуемое Менцером предварительное
утомление, на мой взгляд, также неэффективно.
Менцер исходил из того, что стимулом к росту
является именно отказ мышцы, поэтому считал,
что та мышца, которая откажет первой, и будет
расти, для чего предлагал перед базовым
упражнением утомить нужную мышцу изолированным
движением. Действительно, в свете сделанных
предположений о роли низкого уровня
энергетических фосфатов в разрушении
сократительных структур мышц очень заманчивым
кажется снизить уровень креатинфосфата (а при
возможности — и АТФ) в тренируемой мышце
перед основным базовым движением. Но это не
так просто сделать. Как я уже писал, утомление
мышцы происходит в основном по причине
накоплением кислых продуктов метаболизма в
мышце, что мешает мостикам миозина сцепляться
с актином и генерировать силу, а следовательно,
не даёт развить высокую скорость расходования
энергии и добиться снижения концентрации АТФ.
Повышение кислотности среды в результате работы
мышц — это самозащита организма от излишнего
утомления и истощения запасов АТФ.
Изолированное движение уступает базовому в
скорости расходования АТФ и не может оказать столь
мощного эффекта на мышцу. Перерыв в нагрузке,
связанный с переходом от изолированного
упражнения к базовому, позволит несколько
восстановить уровень энергетических фосфатов
в мышце, сниженный изолированным подходом, а
вот уровень молочной кислоты в мышце при таком
коротком отдыхе не только не снизится, но даже
несколько возрастёт из-за постепенной
активизации гликолиза (максимум закисления
мышцы наблюдается, как правило, уже после
прекращения нагрузки). Таким образом, после
изолированного упражнения мощность, развиваемая
в базовом упражнении, будет значительно ниже,
чем могла бы быть без предварительного
утомления. Так что эффект от "суперсетов"
Менцера достигается, скорее, не благодаря, а
вопреки предварительному утомлению.
Наконец, главное заблуждение Менцера —
это отрицание другого пути в тренировках.
Чуть выше я упомянул о том, что из тупика
адаптации мышц существует два выхода, и
"Супертренинг" лишь один из них, причём самый
радикальный. Другой путь, более сложный в
понимании и применении, как это ни странно,
известен уже достаточно давно — это
циклирование нагрузки.
В тяжёлой атлетике, пауэрлифтинге и ряде других
силовых видов спорта невозможность непрерывного
прогресса является неопровержимым фактом.
Опытным путём установлено, что дабы сдвинуться
с "мёртвой" точки, необходимо отступить,
снизить нагрузку и затем начать новое
наступление на предельный вес. При этом
спортсмены могут позволить себе тренироваться
до шести раз в неделю, не сталкиваясь ни с какой
"перетренированностью" (которая, по мнению
Менцера, неминуемо должна проявиться), а,
наоборот, добиваются нового, более высокого
результата.
Вот как в рамках предложенной выше теории
объясняется необходимость циклирования нагрузки.
Я уже показал во второй части, что сила мышц
зависит не только от содержания в них
сократительных структур, но и от концентрации
энергетических фосфатов, а силовая выносливость
зависит ещё и от способности длительное время
поддерживать концентрацию энергетических
фосфатов на высоком уровне. Достижение
максимального силового результата возможно
только в условиях максимальной реализации
энергетического потенциала мышцы, но тренировки
в таком высокоэнергетическом состоянии
неспособны вызвать во внутренней среде мышц
изменения, достаточно сильные для запуска
дальнейших адаптационных процессов, приводящих
к росту сократительных структур. Поэтому после
достижения пика формы в начале нового
тренировочного цикла нагрузки на мышцы резко
снижают (а иногда практикуют и перерыв в
тренировках), что приводит к некоторой
дезадаптации мышц в энергетическом плане, но
не оказывает существенного воздействия на
сократительные структуры мышц. Последующее
повышение нагрузки вновь оказывает на мышцу
необходимое стрессовое воздействие, что,
соответственно, вызывает рост сократительных
структур мышц. Со временем адаптационные
процессы, происходящие в мышцах, опять
снижают восприимчивость мышц к нагрузке,
получение микротравм блокируется, мышца
достигает пика формы, но для достижения
дальнейшего прогресса требуется вновь
отступить, снизить нагрузку. Это грубая схема,
она не отражает полную картину происходящих
процессов, поэтому к секретам построения
тренировочных циклов я вернусь чуть позже.
А пока в рамках предложенной гипотезы я
попытаюсь ответить на ряд вопросов,
касающихся правил построения тренировок,
направленных на развитие сократительных
структур мышц и основанных на принципах
высокоинтенсивного тренинга с низкой
частотой тренировок, сторонником которого
и являлся автор "Супертренинга".
Тип и темп напряжения мышц
Взять, к примеру, извечный спор на тему: каким
должно быть напряжение мышц? Напряжением
позитивного движения? Или негативного движения?
Или вообще статическим? А каким должен быть
темп движения? Взрывным? Подчёркнуто медленным?
Среди методистов тренинга по этому вопросу не
существует единства мнений. Что и не
удивительно. Если исходить из того, что цель
тренинга — это получение микротравм в результате
снижение концентрации креатинфосфата в мышце до
выравнивания скоростей расходования и ресинтеза
АТФ за счёт основных источников энергии,
становится ясно, что достигнуть эту цель можно при
использовании любого напряжения. Следует иметь
в виду лишь то, что по мере повышения
тренированности мышц добиваться вышеуказанной
ситуации становится всё труднее, поэтому от
тренировки к тренировке скорость расходования
энергии должна возрастать, то есть должна
возрастать средняя мощность подхода. Добиться
увеличения мощности можно не только благодаря
увеличению веса снаряда, но и благодаря
изменению техники движения.
Так, на начальном этапе тренинга достаточно
окажется позитивного движения и свободного
опускания снаряда. При этом энергия будет
расходоваться только при подъёме веса, а
во время обратного движения и отдыха в нижней
и в верхней точках траектории запасы АТФ
начнут восполняться. Искусственное замедление
движения во время негативной фазы приведёт к
тому, что энергия станет расходоваться не
только на подъём снаряда, но и на его
опускание, что увеличит среднюю мощность
подхода. Сокращение времени отдыха между
повторениями (непрерывное движение) ещё больше
увеличит скорость расходования энергии.
Взрывной стиль при подъёме снаряда позволит
использовать больший вес, что в сочетании с
акцентом на негативной фазе ещё больше
повысит скорость расходования энергии (кстати,
именно такой стиль выполнения движения
практикует Дориан Ятс). Но и это не предел
мощности: если значительно повысить вес
снаряда и поднимать его с помощью партнёра,
а опускать самостоятельно (негативные
повторения), то можно добиться максимальной
мощности в подходе. Мышцы новичка будут
изнывать от боли после тренировки с
использованием исключительно позитивного
движения, а опытные регулярно тренирующиеся
атлеты могут получить микротравмы только при
условии использования приёмов по увеличению
средней мощности подхода — например, при
использовании негативных повторений. Этот
факт и позволил распространиться заблуждению,
что микротравмы волокон возникают
исключительно во время негативной фазы
движения. ***
Хотя сам я не исключаю возможность того, что
во время негативной фазы разрывается большее
количество миозиновых мостиков, чем при
позитивном движении. Сие возможно в том
случае, если предел прочности мостика
при растяжении в различных направлениях
различен, а, судя по известной на
сегодня конфигурации молекулы миозина, это
не исключено. Но напоминаю, что причиной
разрыва является не само негативное движение,
а энергетическое состояние мышцы в момент
этого движения.
Количество повторений
Другой извечный вопрос: каково должно быть
количество повторений в упражнении?
Определяющим является не количество повторений,
а время нахождения под нагрузкой. Вероятность
возникновение микротравм существенно возрастает
после снижения концентрации креатинфосфата
в два раза по сравнению с исходным уровнем, что
в быстрых волокнах наступает примерно на
седьмой-десятой секундах максимально
интенсивной работы. В то же время вероятность
получения микротравм значительно снижается
после достижения максимальной скорости
воспроизводства АТФ за счёт гликолиза и
снижения АТФ-азной активности мышц по причине
накопления кислых продуктов метаболизма, что
наблюдается примерно через 30-40 секунд после
начала работы. Таким образом, для достижения
требуемого эффекта в быстрых волокнах подход
должен длиться не меньше семи и не больше
сорока секунд.
Так сколько же времени должна длиться нагрузка? С
одной стороны, интенсивная работа до отказа
длительностью 7 секунд обеспечивает
более высокую скорость расходования энергии, чем
работа длительностью 40 секунд.
Однако малая длительность семисекундной работы
не обеспечивает максимального воздействия на
мышцу. С другой стороны, работа в 40 секунд
длится значительно дольше, причём уже после
существенного снижения уровня креатинфосфата
— но развиваемая при этом скорость расходования
энергии значительно ниже, чем в первом случае.
По-видимому, оптимальная длительность нагрузки
лежит где-то посередине указанного интервала
и составляет 20-30 секунд.
Почему же разные мышцы даже у одного человека
лучше реагируют на разное количество
повторений? Дело в том, что различные
движения длятся разное время. Чем больше
амплитуда движения, тем дольше длится
повторение и тем меньше повторений укладывается
в требуемое время. Так, за 20-30 секунд работы
в таких движениях, как приседания или становая
тяга, можно сделать 6-8 повторений, в жимовых
упражнениях и при работе на бицепс или трицепс
— 8-10 повторений, а вот в упражнениях на
голень и предплечья количество повторений может
составить уже 10-15. Как можно видеть, никакого
принципиального различия в тренировке бицепса,
четырёхглавой мышцы бедра или голени не
существует, поскольку во всех случаях, несмотря
на разное количество повторений, нагрузка длится
примерно одинаковое время.
Но нередко рекомендуемое некоторыми практиками
количество повторений выходит за указанные выше
рамки и достигает 20-30 повторений даже в таких
упражнениях, как приседания и становая тяга.
Всё дело в том, что мышцы состоят не только из
быстрых, но и из медленных волокон, и тренировке
медленных волокон тоже полезно уделять внимание.
Как я уже писал выше, скорость расходования АТФ
в медленных волокнах значительно ниже, чем в
быстрых, поэтому сокращение медленных волокон
энергетически более устойчиво. Создать временный
дефицит АТФ, необходимый для получения
микроразрывов в медленных волокнах, за промежуток
времени в 15-20 секунд практически невозможно.
Но поскольку интенсивность гликолиза в
окислительных волокнах невысока, а максимальная
скорость производства АТФ окислительным путём
может быть достигнута только через 60-120 секунд
после начала работы и напряжение мышцы при
нагрузке свыше 30% от максимальной практически
останавливает кровообращение (что затрудняет
доставку кислорода и получение энергии
окислительным путём), то при предельно
интенсивной работе длительностью 60-90 секунд
можно ожидать достижения дефицита АТФ и
микроразрывов в медленных волокнах.
Размер вклада медленных волокон в общую
гипертрофию мышц зависит от их доли в составе
мышц. По-видимому, утверждения о том, что большое
количество повторений лучше воздействует на
определённые мышцы, исходят от людей, у которых в
данных мышцах преобладают медленные волокна. В
общем случае следует признать, что максимального
развития мышц можно добиться, только уделяя
внимание волокнам всех типов.
Отказ
Следующий очень важный вопрос: нужно ли работать
до отказа, как это утверждает Менцер, и есть ли
какое-то особое магическое значение у последнего
отказного повторения?
Вероятность микроразрывов миофибрилл значительно
повышается, когда после нескольких секунд работы
существенно снижается уровень креатинфосфата в
мышцах. И чем дольше продлится нагрузка в таком
состоянии, тем больше микротравм получит мышца.
Однако по мере активизации основных источников
энергии и падения АТФ-азной активности мышц
скорости расходования и ресинтеза АТФ выравниваются,
и вероятность получения микротравм значительно
снижается. Таким образом, чем дольше длится
подход, тем менее эффективны последние повторения,
а уж работа сверх "отказа" в этом случае
практически бесполезна. Только в том случае,
если отказ наступает в первые 10 секунд работы,
последние повторения играют значительную роль.
Следовательно, при работе в рекомендованном
диапазоне длительности нагрузки 20-30 секунд
последнее отказное повторение по степени
воздействия на мышцы принципиально не отличается
от предыдущих. Следует иметь в виду, что если
изначально цель тренировки будет достигнута при
работе необходимой длительности с относительно
небольшим весом (и, соответственно, без достижения
отказа), то по мере тренированности мышц придётся
повышать вес снаряда. И в конечном счёте всё равно
прийти к тому, что отказ неминуемо наступит в
требуемом диапазоне длительности. Так что я
советую работать до отказа, но помнить, что цель
не в нём, и потому не нужно особо упираться в
последнем повторении в травмоопасных упражнениях,
требующих высокой концентрации внимания и
соблюдения чёткой техники выполнения движения —
таких, как становая тяга или приседания со
штангой на плечах.
Количество подходов и время отдыха между ними
Для того чтобы определить оптимальное количество
повторений и длительность отдыха между ними,
следует разобраться, для чего вообще используется
интервальный метод тренировок — метод чередования
нагрузки и отдыха.
Нагрузки, повторяющиеся через определённый интервал
отдыха, используются для более сильного воздействия
на тренируемую функцию. В зависимости от
особенностей происходящих в организме процессов
можно выделить два принципа взаимодействия нагрузок
в ходе одного тренировочного занятия.
В самом простом случае эффект, достигаемый в ходе
каждого подхода, не зависит от предшествующей
нагрузки. Время отдыха между подходами в этом случае
строго не регламентировано, оно должно быть лишь
достаточным для восстановления сил, дабы атлет имел
возможность повторить последующий подход на
требуемом уровне мощности. Общий эффект от такой
тренировки является простой суммой тренировочных
эффектов, достигнутых в ходе отдельных подходов.
Примером тут может служить тренировка, направленная
на развитие гликолитической ёмкости мышц, срочная
тактическая цель которой — существенное исчерпание
запасов гликогена в мышце, дабы спровоцировать его
сверхвосстановление в период отдыха. В ходе одного
подхода расходуется определённое количество
гликогена, пропорциональное выполненной работе.
Молочная кислота, накапливаемая в мышце в
результате гликолитического способа восстановления
АТФ, останавливает работу задолго до исчерпания
запасов гликогена в мышце. Многократно повторяя
нагрузку после отдыха, достаточного для
существенного вывода молочной кислоты из мышцы,
можно добиться значительного снижения концентрации
гликогена.
При определённых целях тренинга эффект от
последующей нагрузки может не просто линейно
суммироваться с эффектом, полученным в предыдущем
подходе, но и усиливать его. Так, в случае с
короткой интенсивной нагрузкой максимум потребления
кислорода наблюдается не во время самого подхода,
а несколько позже, когда происходит так называемый
"возврат кислородного долга" — восстановление за
счёт окисления макроэнергетических фосфатов (АТФ и
креатинфосфата), израсходованных в ходе интенсивной
работы. Задание повторных нагрузок после
существенного восстановления уровня
макроэнергетических фосфатов, но при сохраняющемся
некоторое время высоком уровне потребления кислорода,
приводит к повышению уровня потребления кислорода от
подхода к подходу, что оказывает более сильный
тренирующий эффект на аэробные способности
организма. В рассматриваемом примере отдых между
подходами уже не может быть произвольным, поскольку
повторная нагрузка после возвращения потребления
кислорода к уровню, соответствующему состоянию
покоя, не даст необходимого тренировочного эффекта.
Скорость восстановления макроэнергетических фосфатов
примерно равна скорости их расходования, поэтому в
рассматриваемом примере отдых после нагрузки по
длительности должен совпадать с длительностью самой
нагрузки — например, 30 секунд
работы, 30 секунд отдыха.
По каким же правилам должны суммироваться
тренировочные эффекты от нагрузок, повторяющихся в
ходе одной тренировки, если цель тренировки —
это разрушение миофибриллярных белков? Как я уже
упоминал, каждый последующий подход по степени
разрушительного воздействия на мышцы менее
эффективен, чем предыдущие, ввиду постепенного
снижения мощности сокращения по причине остаточного
накопления кислых продуктов метаболизма в мышце.
Очевидно, что в этом случае последующая нагрузка
может усилить эффект от предыдущей только за счёт
простого суммирования микротравм, полученных в ходе
каждого отдельного подхода. Следовательно, отдых
между подходами не ограничен какими-либо особыми
условиями, кроме того времени, которым располагает
занимающийся, и должен обеспечивать существенное
снижение концентрации молочной кислоты в мышце,
дабы мышца могла снова развить максимальную
скорость расходования энергии. Полный вывод молочной
кислоты из мышцы при её значительном накоплении
обеспечивается только по истечении нескольких
часов после нагрузки, но для существенного
снижения концентрации молочной кислоты в мышце
достаточно 5-10 минут — для небольших мышц
либо при работе, не связанной со значительным
накоплением молочной кислоты, и 10-20 минут —
для крупных мышечных групп либо при сильном
закислении мышц в ходе подхода. Именно такая
длительность отдыха между подходами обеспечит
максимальный эффект от повторяющихся нагрузок в
рассматриваемом типе тренинга. Короткий интервал
отдыха, который так любят многие бодибилдеры,
обеспечивающий чувство "закачки" мышц, приводит
лишь к максимальному закислению мышц и крови,
что может быть полезно для развития
сопротивляемости организма к снижению рН внутренней
среды, но не имеет прямого отношения к
стимулированию последующего роста мышц.
Итак, со временем отдыха разобрались. Каково же
оптимальное количество подходов?
Как я уже отмечал, молочная кислота не выводится из
мышцы полностью даже при отдыхе между подходами
в 10-20 минут, то есть развиваемая мощность
сокращения в каждом последующем подходе будет
несколько ниже, чем в предыдущем. Задавать
повторную нагрузку на мышцу имеет смысл только на
определённом требуемом уровне интенсивности,
поэтому после существенного снижения мощности мышц,
развиваемой во время подхода, нагрузку на данную
мышцу следует прекращать. Эксперименты показывают,
что переломный момент в развиваемой мощности
наступает в среднем после пятого-шестого подхода
в упражнении. По-видимому, именно это количество
подходов для тренировки одной мышечной группы и
следует признать оптимальным в рассматриваемом
режиме тренировки.
Данное количество повторений оптимально для
максимального разрушения миофибриллярных белков,
но является ли максимальное разрушение оптимальным
для достижения максимального сверхвосстановления
мышц во время отдыха?
Закон восстановления энергетических резервов
гласит, что чем больше расход энергии при работе
мышц, тем интенсивнее протекают процессы
восстановления и тем значительнее превышение
исходного уровня энергетических ресурсов в фазе
сверхвосстановления. Однако при чрезмерно
интенсивной работе, связанной со значительным
накоплением продуктов метаболизма, скорость
восстановительных процессов может снизиться, а
фаза сверхвосстановления будет достигнута в
более поздние сроки и выражена в меньшей степени.
По-видимому, этому же закону подчиняются и
процессы восстановления белковых структур мышц.
Как я уже показал во второй части, чрезмерные
разрушения затрудняют процессы восстановления и
могут привести даже к отрицательному результату.
Поэтому следует признать, что количество
микротравм, полученных в ходе тренировки, должно
быть не максимальным, а оптимальным, достаточным
для того, чтобы инициировать восстановительные
процессы, но не таким большим, чтобы сорвать
восстановительные возможности организма.
Однозначно указать количество подходов,
необходимое для достижения оптимального
количества микротравм, невозможно, поскольку это
количество зависит от уровня тренированности
мышц и интенсивности задаваемой нагрузки.
Так, даже один интенсивный подход (здесь и далее
в виду имеется интенсивность, позволяющая
выполнять упражнение в рамках необходимого
диапазона длительности нагрузки) может быть
эффективней нескольких менее интенсивных
подходов, а несколько высокоинтенсивных подходов
могут оказаться слишком разрушительными для
организма.
В регулировании уровня тренировочной нагрузки
существует два конкурирующих методических
подхода.
Первый — когда объём выполняемой нагрузки
задаётся заранее (например, 5-6 подходов),
но на заданном уровне интенсивности и при заданной
длительности подхода (заранее известном весе
снаряда и количестве повторений). Подходы тут,
естественно, выполняются не до отказа, а
прерываются по выполнении заданной работы. По
мере повышения тренированности мышц повышается
и интенсивность подходов (вес снаряда), и таким
способом осуществляется чёткое дозирование
нагрузки. Ошибкой в данном случае является
стремление не регулировать нагрузку заранее, а
выполнять все подходы до отказа на пределе
интенсивности, стараясь выжать из организма
максимум.
Второй подход основывается на диаметрально
противоположном принципе — в работе используется
нагрузка максимальной интенсивности, которую
можно развить в рамках необходимого диапазона
длительности, а вот общее воздействие тренировки
на мышцу регулируется количеством повторений
такой нагрузки (то есть количеством подходов). В
этом случае необходимое количество подходов, как
правило, оказывается меньшим, чем в первом методе.
Достигаемый эффект зависит не только от величины
нагрузки, но и от уровня тренированности мышц.
Так, в нетренированных мышцах даже один подход
далеко не предельной интенсивности вызывает
сильнейшие разрушения (пусть читатель вспомнит
свои ощущения на следующий день после первого
посещения тренажёрного зала) и, наоборот, в
тренированной мышце даже множество
высокоинтенсивных подходов может не вызвать
необходимого эффекта.
Как отмечалось, при редких тренировках,
рекомендуемых Менцером, энергетический потенциал
мышц остаётся на довольно низком уровне даже в
течение длительного периода тренировок, поскольку
срочный тренировочный эффект в энергетической
сфере не переходит в долговременную адаптацию
ввиду большого перерыва в тренировках — что и
облегчает воздействие нагрузки на мышцу. В
"Супертренинге" Менцера каждая тренировка по
разрушающему миофибриллярные белки эффекту близка
к "первой" тренировке — и именно поэтому при
таком виде тренинга оказывается достаточным
одного-единственного "отказного" подхода.
Подбор упражнений
Итак, если для роста мышцы может быть достаточно
даже одного подхода, то кажется, что тренировать
мышцу несколькими упражнениями тем более ни к
чему. С другой стороны, варьируя используемые
упражнения, можно добиться воздействия на
различные пучки мышечных волокон, что будет
способствовать развитию пропорциональной
мускулатуры. Однако тяжёлый тренинг, основанный
на стимулировании роста мышц путём их
предварительного разрушения, требует напряжения
восстановительных функций организма, и когда
последнему придётся делить ограниченные
пластические и энергетические ресурсы между
всеми мышцами, нуждающимися в восстановлении, то
результат восстановления может разочаровать
занимающегося. Каждому бодибилдеру приходится
искать баланс между желанием получить идеальную
фигуру и восстановительными резервами организма.
Если атлет не использует в период тяжёлых
тренировок дополнительные "восстановители",
то лучше остановить выбор на нескольких крупных
мышечных группах и базовых упражнениях и не
распылять силы на весь спектр существующих
движений — тем более, что влияние
упражнений на форму мышц сильно преувеличено. То,
каким будет бицепс, определяется генетически, а
не тем, что атлет делает на скамье Скотта:
подъёмы гантелей или концентрированные сгибания.
На юношеских фотографиях Шварценеггера хорошо
просматриваются знакомые очертания мышц будущего
Мистера Олимпия, то есть годы тренировок
практически не изменили форму мышц Арнольда, а
лишь увеличили их.
Почему я настаиваю именно на базовых упражнениях
со свободными весами, а не рекомендую
воспользоваться тем или иным тренажёром? Кажется,
что, с точки зрения теории, атлет может
расходовать энергию любым способом. То есть
делает ли атлет упражнения в тренажёре или с
помощью штанги — это якобы не имеет значения.
Но так кажется только на первый взгляд. Большинство
тренажёров основаны на системе блоков, сила трения
в которых достигает порой значительных величин. В
результате позитивное движение в тренажёрах
затруднено, но зато при обратном (негативном)
движении (а именно в негативном движении мышца
способна развить максимальную силу, достичь
максимального напряжения) сила трения облегчает
работу — в результате чего средняя мощность
подхода в тренажёре ниже, чем в аналогичном
движении со свободным весом, что, естественно,
отрицательно сказывается на тренировочном эффекте.
Из существующих тренажёров могу порекомендовать
только тренажёры рычажного типа с навешивающимися
дисками, — работа в них аналогична работе со
свободными весами. Отмечу также, что тот, кто
изобретёт тренажёр, в котором опускать вес будет
тяжелее, чем поднимать его, произведёт переворот
в бодибилдинге.
Частота тренировок
Перехожу к главному вопросу, вызывающему
наибольшее количество споров: какова должна быть
длительность отдыха между тренировками?
Рекомендую обратиться ко второй части данного текста
и вспомнить, что в самом простом случае величина
отдыха между тренировками определяется временем,
необходимым для восстановления и достижения
состояния сверхвосстановления, суперкомпенсации
ведущей тренируемой функции.
Ещё совсем недавно было распространено убеждение,
что для восстановления мышцы после тренировки
достаточно сорока восьми часов. Возможно, в этом
убеждении есть доля истины, поскольку примерно
через такой период отдыха и наблюдается
суперкомпенсация параметров, ответственных за
энергетический потенциал мышцы. В случае же
получения микротравм через 48 часов мышца не
только не восстановится, но даже не успеет
очиститься от повреждённых структур. Тогда,
может быть, для восстановления мышц требуется
5-7 дней? Именно к этому сроку обычно полностью
проходят болевые ощущения в мышцах после
тяжёлой нагрузки, и именно такой период отдыха
становится популярным в последнее время.
Вынужден разочаровать читателей: исчезновение
болевых ощущений вовсе не означает, что мышца
восстановилась и что достигнут эффект от
тренировки. Исчезновение болевых ощущений
означает лишь то, что в мышце закончилось
воспаление, сопровождающее процессы лизиса
повреждённых структур. Для полного восстановления
повреждённых волокон и достижения состояния
"суперкомпенсации" в зависимости от величины
повреждений может потребоваться ещё не менее
недели. Получается, что тренировка,
сопровождающаяся микротравмами мышц и сильными
болевыми ощущениями, не должна практиковаться
чаще, чем один-два раза в месяц на одну группу
мышц. Применение стероидов может сократить
время отдыха, но не кардинально. Стероиды делают
фазу суперкомпенсации просто более выраженной.
Предвижу следующие возражения: если для
восстановления после тренировки требуется около
двух недель, то как же удаётся получить рост
мышечной массы при тренировках одной мышцы
два-три раза в неделю? Дело вот в чём: плохо ли,
хорошо ли, но мышца растёт и в этом случае. На
начальном этапе таких занятий мышечные волокна
повреждаются на каждой тренировке, и ни о каком
полноценном восстановлении речь не идёт. Одни
микротравмы накладываются на другие, и так
продолжается около месяца до тех пор, пока
значительно не возрастает энергетический
потенциал мышцы, что блокирует получение
микротравм, и только после этого в мышцах
начинают преобладать восстановительные процессы.
Таким образом, при частых тренировках
гипертрофия мышц становится возможной только
после существенной адаптации мышц к задаваемой
нагрузке. Процессы восстановления и роста длятся
ещё около месяца, и на этом, если ничего не
менять в тренировках, рост мышечной массы и
силовых показателей заканчивается — по причине
всё той же адаптации мышц к нагрузке и
отсутствия нового стимула к росту. Как правило,
требуется ещё около двух месяцев на то, чтобы
понять, что выбранная методика тренировок
перестала давать результаты и попытаться что-то
изменить в тренировках.
Итак, на достижение гипертрофии мышц при обычных
тренировках (наиболее распространённых по всем
тренажёрным залам) требуется два-три месяца.
Практически такие же результаты в увеличении
мышечной массы (но не в увеличении
работоспособности) можно получить от нескольких
тренировок, давая мышцам полноценный отдых
длительностью полторы-две недели, а не дожидаясь,
пока мышцы добьются отдыха сами, адаптировавшись
к нагрузке.
Что в итоге?
Из сделанных теоретических предпосылок в качестве
системы тренировок, направленной на развитие
сократительных структур мышц, постепенно
вырисовывается система, сильно напоминающая
"Супертренинг" Менцера, отличительными чертами
которой являются:
— ограниченное количество используемых упражнений
(одно-два базовых упражнения на одну мышечную
группу);
— ограниченное количество высокоинтенсивных
"отказных" подхода в каждом упражнении;
— длительный отдых между тренировками одной
мышечной группы (полторы-две недели).
Казалось бы, эти положения полностью противоречат
принципам современного бодибилдинга, основой
которого являются практически ежедневные,
высокообъёмные тренировки. Критики системы
Менцера утверждают, что данная система
противоречит основам теории физической культуры и
вообще физиологии человека и не может дать
никакого результата. Однако как я показал выше,
это не так. Просто для адептов классического
тренинга рост мышц стал неразрывно связан с
объёмом выполняемой работы. Между тем, как я
показал выше, стимулом к росту мышечной ткани
является не собственно объём работы, а изменения
внутренней среды мышц, возникающие в ходе
тренировки. Объём работы лишь средство,
вызывающее упомянутые изменения, но далеко
не единственное, — повышение интенсивности
выполняемой работы оказывает более значительное
влияние на состояние внутренней среды мышц даже
при незначительном объёме выполняемой работы.
Отдых же между тренировками в "Супертренинге"
определяется временем наступления
сверхвосстановления, суперкомпенсации ведущей
тренируемой функции — количества
сократительных структур мышцы. Развитие всех
остальных параметров, имеющих меньшее время
достижения фазы суперкомпенсации (например,
количество гликогена, креатинфосфата, окислительных
ферментов, ферментов гликолиза в мышце и др.),
попросту игнорируется — что, конечно,
сказывается на объёме и силе мышц, но облегчает
воздействие тренировки на мышцы и практически
избавляет от такой проблемы бодибилдинга, как
излишняя адаптация мышц к нагрузке. А длительный
период отдыха обеспечивает полноценное
восстановление и избавляет от другой возможной
проблемы — перетренированности.
Не следует ожидать от "Супертренинга" развития
тех качеств, для тренировки которых он не
предназначен. Вклад сократительных структур в
силу и массу мышц является наиболее значительным,
но далеко не единственным. Соответственно, при
всех его достоинствах "Супертренинг" не может
обеспечить максимально возможное развитие силы и
массы мышц.
Наиболее полное развитие мышц и силовых качеств
спортсмена могут обеспечить лишь разносторонние
тренировки, направленные на улучшение всех
основных двигательных функций и на рост всех
основных компонентов мышечного волокна.
Прежде чем приступить к рассмотрению правил
построения таких многоцелевых тренировок,
следует систематизировать тренировочные цели, а
затем определить основные функции, развитие
которых может привести к достижению поставленных
целей.
Часть IV. Основные цели
тренинга
В зависимости от специализации спортсмена в
качестве основной цели тренинга можно выделить
развитие следующих качеств мышц:
— сила, развиваемая мышцами в специализированных
движениях (пауэрлифтинг, тяжёлая атлетика);
— силовая выносливость (гиревой спорт, борьба,
бег на 200 м и на 400 м);
— мышечные объёмы (бодибилдинг).
Сила
Для начала нужно разобраться, от чего зависит
результат, достигаемый спортсменом в
специализированных движениях тяжёлой атлетики
или пауэрлифтинга. Проявление скоростно-силовых
качеств мышц в упомянутых видах спорта несколько
различны. Цель пауэрлифтёра — поднять
максимальный вес вне зависимости от скорости
движения. В тяжёлой же атлетике соревновательные
движения технически более сложны и конечный
результат зависит от того, будет ли штанга в
нужной точке траектории иметь нужную скорость
движения. Масса снаряда в тяжёлой атлетике
относительно ниже, чем в пауэрлифтинге, однако
снаряд приходится разгонять до значительно
бо́льших скоростей. Но при внимательном
рассмотрении различия между тяжёлой атлетикой и
пауэрлифтингом оказываются не столь уж и
существенными. Дело в том, что сила, которую
спортсмену необходимо прикладывать к снаряду для
его равномерного подъёма (вернее, проекция силы
на вертикальную ось), равна произведению массы
снаряда на ускорение свободного падения (напоминаю
второй закон Ньютона: F = mg (буквами F принято
обозначать силу, m — массу, а g —
ускорение свободного падения)). Конечно, в
момент отрыва для придания снаряду начальной
скорости необходимо приложить несколько бОльшую
силу, поскольку снаряду необходимо сообщить
начальное ускорение. Сила, которую необходимо в
этом случае приложить к снаряду, равна F = m(g + a),
где а — ускорение, сообщаемое снаряду. Различие
между пауэрлифтингом и тяжёлой атлетикой проявляется
как раз в величине этого ускорения. В пауэрлифтинге
штанге необходимо сообщить лишь минимальное
ускорение, достаточное для сдвигания её с места и
придания минимальной скорости, достаточной для
прохождения "мёртвой" точки. В тяжёлой же атлетике
требование к развиваемому ускорению значительно
выше, чем в пауэрлифтинге. Но и в том и в другом
случаях результат зависит от силы, прикладываемой
к снаряду. Чем выше сила, тем больше масса штанги,
которой пауэрлифтёр может придать минимально
необходимое ускорение, и тем больше ускорение,
которое тяжелоатлет может придать штанге с
определённой массой. Таким образом, в обоих этих
видах спорта результат зависит от силы,
прикладываемой к снаряду, а соответственно, он
зависит от силы, развиваемой мышцами.
Впрочем, между пауэрлифтингом и тяжёлой атлетикой
можно найти ещё одно различие: сила мышц зависит
от скорости их сокращения (вспомните соотношение
Хилла, упомянутое мной в первой части: чем выше
скорость сокращения мышцы, тем меньшую силу она
способна развить). В тяжёлой атлетике мышцы
вынуждены сокращаться при значительно бо́льших
скоростях, чем в пауэрлифтинге, поэтому и
развиваемая ими сила заметно меньше.
Результат в соревновательном движении зависит
не только от силы мышц, но и от оптимальной
траектории движения, а также от своевременного и
эффективного приложения силы к снаряду, то есть
от техники выполнения упражнения. Я не буду
подробно останавливаться на этом вопросе, поскольку
не считаю себя в нём достаточно компетентным, тем
более что за годы развития таких видов спорта,
как тяжёлая атлетика и пауэрлифтинг, накоплен
богатый методический материал по постановке техники
соревновательных движений. Отмечу только, что
основой освоения техники выполнения упражнения
является выполнение большого количества
однотипных движений, что приводит к закреплению
в центральной нервной системе двигательного
стереотипа. Видимо, отчасти по этой причине
одним из существенных критериев в планировании
тренировочных нагрузок на начальном этапе
тренинга в классической спортивной школе
является показатель КПШ — количество
подъёмов штанги.
Давайте лучше рассмотрим, от чего зависит
собственно сила, развиваемая мышцами, и какие
методы тренировки могут повлиять на способность
мышц генерировать силу. Как я отмечал в первой
части, сократительным элементом волокна является
миофибрильная нить. Силу, развиваемую миофибриллой,
генерируют боковые выступы молекулы миозина,
называемые мостиками, которые совершают гребковые
движения. Обращаю внимание читателя на тот факт, что
миофибрилла — это цепочка последовательно
соединённых саркомеров, а крепость цепи, как
известно, зависит от крепости любого её звена. Сила
миофибриллы как целого не может быть больше силы её
части — отдельного саркомера. То есть каждый
саркомер должен развивать одинаковую силу, и эта
сила равна силе всей миофибриллы. Сила, развиваемая
саркомером, зависит от его длины — то есть чем
длиннее саркомер, тем бОльшим количеством
миозиновых мостиков он располагает и тем сильнее его
сокращение. Мышечные волокна разных мышц и даже
одних и тех же мышц, но у различных индивидов, имеют
разные длины саркомеров и, соответственно, разную
способность к генерации силы. Однако длина саркомера
задаётся генетически и не поддаётся тренировке,
поэтому в дальнейшем влияние длины саркомера на силу
я даже не буду рассматривать.
Из вышеизложенного можно сделать вывод, что сила
мышцы зависит не от длины миофибрильных нитей (от
этого зависит амплитуда сокращения мышцы), а от
количества миофибрильных нитей в поперечном сечении
мышцы. И вот этот параметр как раз поддаётся
развитию.
Основные принципы тренировки, нацеленной на рост
сократительных структур мышц, я рассматривал во
второй части. Напомню основную фабулу.
Высокоинтенсивные тренировки, приводящие к
сокращению мышц в условиях недостатка
макроэнергетических фосфатов, разрушают
сократительные белки мышечных волокон.
Микротравмы мышечных волокон запускают
восстановительные процессы, приводящие к
делению клеток-сателлитов и к увеличению
числа клеточных ядер в мышечных волокнах,
что при условии достаточно длительного и
полноценного восстановления приводит к
увеличению сократительных структур в мышце.
Я также показал, что сила, развиваемая мышечным
волокном, и скорость его сокращения зависят от
насыщенности волокна АТФ. Поскольку сокращение
мышц не мгновенно, поскольку оно длится некоторое
время даже при единичных повторениях упражнения и
сопровождается расходом АТФ, то результат
выполнения упражнения зависит ещё и от способности
мышц быстро восстанавливать уровень АТФ,
то есть от концентрации в волокне креатинфосфата и
креатинкиназы.
Содержание креатинфосфата в мышцах спортсменов
в 1,5-2 раза выше, чем у нетренированных людей.
Соответственно, данное качество мышц поддаётся
тренировке. Какой же вид тренировки наиболее
эффективен для целей повышения в мышцах концентрации
креатинфосфата?
Содержание креатина в мышцах значительно превышает
концентрацию собственно креатинфосфата. Так, общая
концентрация креатина в мышцах составляет в среднем
120 ммоль/кг, в то время как с фосфатом связано (то
есть является креатинфосфатом) только
около 70 ммоль/кг. Таким образом, существенная часть
креатина в мышцах находится в не связанном с
фосфатом состоянии, и резерв увеличения
концентрации креатинфосфата заключается как раз в
этом не связанном с фосфатом креатине: необходимо
лишь заставить мышцы фосфорилировать больше креатина.
Существенное снижение концентрации креатинфосфата
во время интенсивного сокращения мышц (то есть
отсоединение от него фосфата и превращение просто
в креатин) сразу по прекращении работы приводит к
интенсификации процессов, восстанавливающих его
уровень.
Во время отдыха, благодаря окислению, АДФ
и фосфат, в избытке накопившиеся в мышце в
результате гидролиза АТФ при работе миозиновых
мостиков и кальциевых насосов, вновь превращаются
в АТФ, а затем фосфатная группа переносится
с АТФ на креатин с образованием
креатинфосфата. В результате концентрация креатинфосфата
в мышце уже через несколько минут отдыха не только
восстанавливается, но и превышает исходный уровень,
характерный для состояния покоя. То есть
наблюдается сверхвосстановление креатинфосфата в
мышце. Однако такое состояние длится недолго, и
концентрация креатинфосфата снижается уже через
пару часов. Проводя повторные нагрузки на мышцу в
состоянии сверхвосстановления, то есть после
отдыха в несколько минут, можно добиться заметного
повышения концентрации креатинфосфата. Правда, уже
через несколько часов концентрация последнего
существенно снижается, но, по-видимому, некоторое
превышение исходного уровня сохраняется дольше,
поскольку регулярные тренировки (не реже двух-трёх
раз в неделю) приводят к постепенному относительно
стойкому повышению концентрации креатинфосфата в
мышцах. В противовес этому перерыв в тренировках
дольше, чем на одну неделю, заметно снижает
уровень креатинфосфата.
Рассмотрю чуть подробнее принципы тренировок,
направленных на развитие креатинфосфатной мощности
и ёмкости мышц. Уровень нагрузки при таких
тренировках должен быть достаточно высоким (чтобы
активировать бОльшую часть мышечных волокон и
обеспечить высокую скорость расходования
энергии) и составлять 70-85% от единичного
максимума.
Длительность нагрузки должна быть таковой, чтобы
запасы креатинфосфата в мышце были использованы
не менее чем наполовину, то есть нагрузка должна
длиться не меньше семи секунд. В то же время
работу желательно прекращать до активации
гликолиза, поскольку накопление молочной кислоты
в мышцах приводит к замедлению темпов
восстановления АТФ и креатинфосфата.
Соответственно, стремиться к полному отказу мышц
не следует, и нагрузка не должна длиться дольше
пятнадцати секунд. Соответственно, количество
повторений в подходе должно быть в районе 4-6.
Отдых между подходами должен длиться около
3-5 минут, что необходимо для обеспечения
сверхвосстановления уровня креатинфосфата. И
хотя возможен и более длительный отдых, поскольку
сверхвосстановление длится полтора-два
часа, но, исходя из принципа экономии тренировочного
времени, можно ограничиться тремя-пятью минутами.
Количество таких подходов должно составлять
от 5 до 10, больше
напрягаться просто нет смысла, поскольку
резервы подъёма уровня креатинфосфата в ходе
одного занятия не беспредельны, а вот
усталость будет накапливаться от подхода к
подходу.
Известный тренер по пауэрлифтингу Б.И.Шейко
иногда практикует на подопечных выполнение
двух серий подходов одного упражнения за одну
тренировку. Например, после 5-6 подходов
в жиме лёжа следует нагрузка на ноги, а затем спортсмен
возвращается к выполнению жима лёжа и делает ещё
5-6 подходов. Не знаю, какой смысл Б.И.Шейко
вкладывает в эти действия (возможно, он просто
стремится к общему увеличению объёма нагрузки на
требуемом уровне интенсивности), но, помимо всего
прочего, такого рода практика должна
способствовать повышению уровня креатинфосфата в
мышцах, поскольку возвращение к выполняемому
упражнению после получасового-часового
отдыха происходит на фоне повышенного предыдущими
подходами уровня креатинфосфата.
Рассказывая о методах повышения концентрации
креатинфосфата в мышцах, нельзя не поднять вопрос
об эффективности приёма креатина в качестве
пищевой добавки. Запасы креатина в организме
пополняются благодаря его синтезу в печени, а
также благодаря поступлению креатина с пищей
(с мясными продуктами).
Эксперименты (Harris et al.) показали, что приём
высоких доз креатина (5 г 4-5 раз
в сутки) (5 г креатина эквивалентны
одному килограмму сырого мяса) в течение недели
приводит к существенному увеличению как концентрации
креатина в мышцах, так и концентрации креатинфосфата.
Но наиболее выражен прирост этих показателей при
ежедневных тренировках. Так, содержание креатина в
мышцах в среднем увеличилось с 118,1 ммоль/кг
до 148,5 ммоль/кг в неупражнявшейся мышце, и
до 162,2 ммоль/кг в упражнявшейся. Содержание
креатинфосфата за этот же период возросло
от 81,6 ммоль/кг до 93,8 ммоль/кг
в неупражнявшейся мышце и до 103,1 ммоль/кг
в упражнявшейся мышце. Дальнейший приём креатина не
привёл к существенным изменениям концентрации креатина
и креатинфосфата в мышцах.
Интересно отметить, что некоторые спортсмены,
несмотря на потребление креатина, не получили
существенного прироста вышеуказанных показателей.
Как оказалось, эти спортсмены уже изначально
обладали высокими показателями содержания креатина
в мышцах. В данных экспериментах было убедительно
доказано, что приём сверхдоз креатина с пищей
положительно сказывается на креатинфосфатной
ёмкости мышц, однако о побочных эффектах таких
дозировок ничего не сообщается.
Итак, я рассмотрел методы тренировок,
способствующие развитию силы собственно мышечных
волокон. Сила же мышцы как целого зависит от
того, насколько много волокон одновременно
включены в работу, и от того, с какой частотой
мышечные волокна стимулируются нервными импульсами
(чем выше частота этих импульсов, тем сильнее
сокращение мышцы). Что, в свою очередь, зависит
от того, насколько сильно поляризуется мембрана
тела мотонейрона, расположенного в спинном мозге
под воздействием сигнала, поступающего по сети
нейронов из вышележащих отделов центральной
нервной системы (ЦНС). Путь нервного импульса
начинается в двигательных центрах головного мозга
и проходит по спинному мозгу к мотонейронам,
иннервирующим волокна той или иной мышцы.
Напоминаю, что каждый мотонейрон имеет свой порог
возбудимости и включается в работу только в том
случае, если возбуждение его мембраны превышает
упомянутый порог. Таким образом, чем сильнее
импульс, поступающий от мозга, тем больше
мотонейронов и, соответственно, иннервируемых ими
волокон подключаются к сокращению. Кроме того,
чем сильнее поляризация мембраны мотонейрона, тем
выше частота потенциала действия, возникающего в
мотонейроне, и передающегося по аксону к мышечным
волокнам.
Управление движением — процесс сложный и
запутанный, учёные ещё не до конца в нём
разобрались, а уж я тем более далёк от полного
понимания этого процесса. Поэтому, не влезая в
дебри, опишу лишь его ключевые моменты.
Судя по всему, управление двигательной активностью
организовано так, что мозгу очень тяжело заставить
сокращаться все двигательные единицы (мотонейроны
и иннервируемые ими волокна) одновременно. ЦНС не
генерирует максимальный импульс сразу, а запускает
пробный импульс определённой величины (в
зависимости от ожидаемой нагрузки), который
активирует небольшое количество мотонейронов.
Специальные рецепторы, расположенные в мышцах
(мышечные веретёна), сигнализируют в мозг об
изменениях длины мышцы под действием поступившего
сигнала, и если сокращение не происходит или
скорость его недостаточна (нагрузка слишком
велика), то мозг усиливает запускающий сигнал и
вовлекает в работу большее количество
мотонейронов, одновременно усиливая частоту
потенциала действия уже работающих мотонейронов.
В результате одни волокна вовлекаются в работу
чуть раньше, другие чуть позже, и, таким образом,
максимумы сокращения различных волокон не
совпадают, и двигательные единицы работают
асинхронно (как поршни в двигателе автомобиля).
Таким манером достигается плавность движения, но
не реализуется максимум силы, который мог бы быть
достигнут при совпадении по времени максимумов
сокращения всех волокон мышцы. Между тем
способность к быстрому вовлечению в работу
максимального количества волокон поддаётся
тренировке. Задача атлета — научить мозг
генерировать как можно более мощный запускающий
импульс. Похоже, что развитие таких способностей
подчиняется тем же правилам, что и тренировка всех
иных функциональных качеств спортсмена,
рассматриваемых выше в данном тексте. Прохождение
максимально мощного нервного импульса по всей
цепочке, от двигательных отделов головного мозга
до мышечных волокон, вызывает напряжение всех
элементов этой цепи и ослабление их
функциональных возможностей. То есть наблюдается
физическая усталость — торможение нервной
системы, что выражается в потере способности ЦНС
генерировать и передавать сигнал требуемой силы.
Восстановление функции нервной системы в период
отдыха приводит к сверхвосстановлению её
функциональных возможностей, а регулярное
повторение этих процессов приводит к закреплению
долговременных адаптационных изменений в ЦНС
спортсмена.
Итак, тактическая цель нервно-моторной тренировки
— заставить ЦНС генерировать максимально мощный
нервный импульс. Для чего можно использовать либо
работу с околопредельными весами на 1-3 повторения,
либо работу с умеренными весами, но во взрывном
стиле, то есть атлет должен стараться разгонять
снаряд до максимальных скоростей, прикладывая к
нему по всей траектории максимальную силу. Отдых
между подходами на такой тренировке должен быть
достаточно длительным для восстановления
способности ЦНС и собственно мышц развить
необходимое усилие (то есть отдых должен длиться
5 минут и более — в зависимости от упражнения).
В литературе я не встречал конкретных сведений о
сроках сверхвосстановления возможностей ЦНС после
тяжёлой тренировки, поэтому делать выводы о
необходимом отдыхе между такого рода тренировками
я могу, только исходя из практики силовых видов
спорта. Как правило, серьёзная нагрузка на ЦНС
не практикуется чаще двух раз в неделю, и реже,
чем раз в 7-10 дней.
Но оказывается, что мощный импульс от ЦНС — это
ещё не залог максимальной активации мотонейронов.
Дело в том, что в сухожилиях расположены
специальные рецепторы, так называемые
"органы Гольджи", цель которых — контроль
величины напряжения мышцы. При превышении
напряжения в сухожилиях некоего порога, органы
Гольджи оказывают на мотонейроны мышцы тормозящее
воздействие. Понятно, что благодаря такому
механизму мышца защищает себя от разрывов при
чрезмерной нагрузке. Однако сухожильные рецепторы
не могут точно определить величину критического
напряжения и срабатывают, как правило, с большим
запасом, активизируясь, когда напряжение
значительно превышает привычное. Поэтому цель
спортсмена, стремящегося к поднятию действительно
больших весов — отодвинуть этот защитный барьер.
Один из способов такого воздействия на защитные
механизмы — привыкание сухожилий и рецепторов к
околопредельной нагрузке. Чему может
способствовать всё та же работа с максимальными
весами в 1-3 повторениях, и даже, более
того, — выполнение частичных повторений с
нагрузкой, превышающей единичный максимум, то есть
выполнение полуприседов, тяг с возвышения,
дожимов штанги и пр. Вышеизложенное ещё раз
подтверждает хорошо известный в теории физической
культуры принцип специфичности, который можно
выразить простыми словами: "Что тренируешь, то
и получаешь".
Часть V. Выносливость
Чуть раньше я рассмотрел факторы, от которых
зависит сила, развиваемая мышцами, и методы
тренировок, направленных на развитие силы за
счёт этих факторов. Теперь настало время
разобраться, от чего зависит способность мышц
удерживать необходимый уровень силы
определённое время, то есть от чего зависит
выносливость спортсмена и какие методы тренировок
приводят к развитию общей и специфической
выносливости.
Интервальные тренировки
После возрождения Олимпийских игр в конце XIX века
господствующим методом тренировки на выносливость
стал метод непрерывной работы. Предполагалось, что
интенсивность и продолжительность тренировки должна
соответствовать условиям предстоящих соревнований.
Например, бегуны совершали забеги, равные
соответствующим соревновательным дистанциям,
пытаясь раз от разу улучшить результат в забеге.
В двадцатые годы прошлого века на смену непрерывной
нагрузке пришёл метод интервальной тренировки,
успешное внедрение которого связано с именами
выдающегося финского бегуна П.Нурми и
известного теоретика спортивной тренировки
М.Пикхала. Этими специалистами было показано, что
многократное повторение коротких, но более
интенсивных нагрузок даёт гораздо больший
тренировочный эффект, чем более длительная, но
менее интенсивная работа. В последующие годы
данный тезис получил всё больше и больше
практических подтверждений, а исследователи выявили
биохимические факторы, лежащие в основе
эффективности интервальных тренировок.
Так в чём же преимущество интервальных тренировок?
Для ответа на этот вопрос необходимо
систематизировать множество факторов, влияющих на
работоспособность спортсмена.
Среди факторов, ограничивающих работоспособность,
можно выделить факторы общей выносливости,
определяющиеся возможностями различных систем
организма обеспечивать работу мышц, и
специфические факторы, ответственные за
работоспособность собственно мышц спортсмена.
Общая выносливость лимитируется в основном
способностью организма спортсмена обеспечить
потребность мышц в кислороде и питательных
веществах, а также способностью отводить от мышц
метаболические факторы утомления — такие,
как молочная и угольная кислоты. Напоминаю, что
молочная кислота — это конечный продукт
гликолиза, а угольная кислота получается при
растворении углекислого газа, образующегося в
ходе окисления органических веществ. Таким
образом, общая выносливость определяется
возможностями кровеносной и дыхательной систем
организма, а также запасами органического топлива
(в основном глюкозы в мышцах и печени, а кроме
того, жирных кислот в жировой ткани) и
эффективностью мобилизации топлива в случае
необходимости.
Способность организма поглощать кислород и
выводить углекислый газ зависит прежде всего
от дыхательного объёма лёгких и скорости
газообмена в них.
Возможности кровеносной системы по переносу
кислорода лимитируются общим объёмом крови,
концентрацией в крови гемоглобина (белка-переносчика
кислорода) и скоростью циркуляции крови. Последняя
зависит от ударного объёма сердца (то есть от
объёма крови, прокачиваемого сердцем за одно
сокращение).
Возможности кровеносной системы по отводу кислых
продуктов метаболизма от работающих мышц
определяются, помимо общего объёма крови и скорости
её циркуляции, способностью организма поддерживать
физиологически нормальный уровень рН крови,
скоростью утилизации молочной кислоты и скоростью
вывода углекислого газа через лёгкие. Протекание
многих жизненно важных химических процессов в
организме зависит от кислотно-щелочного
равновесия (рН) среды. Примером может служить
угнетающее влияние повышения кислотности мышечной
саркоплазмы на активность АТФазы миозина,
о котором я рассказывал выше. В состоянии покоя
кислотно-щелочное равновесие крови слегка смещено
в щелочную сторону, и рН крови
составляет 7,4 (в нейтральной
среде рН = 7). Интенсивная мышечная
деятельность сопровождается образованием большого
количества молочной кислоты в мышцах, эта кислота
выводится в кровь, что повышает кислотность крови
и снижает рН до 6,9-6,8.
Организм человека способен выдержать лишь
незначительное снижение рН крови. Так,
в состоянии изнеможения рН может
опуститься до 6,5, при этом наблюдается
тошнота и головокружение. Борьбу с повышением кислотности
крови организм ведёт с помощью буферных реакций.
Вещества, называемые бикарбонатными буферами и
содержащиеся в крови (примером может служить
NaHCO3), вступают в реакцию с молочной
кислотой, образуя соль молочной кислоты и более
слабую угольную кислоту, которая легко распадается
на воду и углекислый газ. Последний выводится через
лёгкие в выдыхаемый воздух, образуя так называемый
неметаболический избыток углекислого газа.
Определяя соотношение вдыхаемого кислорода и
выдыхаемого углекислого газа, можно судить об
интенсивности гликолиза в мышцах.
Зависит рН среды и от скорости вывода молочной
кислоты из крови. Молочная кислота заканчивает
свой метаболический путь либо в сердечной мышце,
где окисляется в митохондриях и служит источником
АТФ для сокращения миокарда, либо в печени, где с
затратой энергии преобразуется обратно в глюкозу
и далее в гликоген, который снова может служить
источником энергии.
Какого же рода тренировки способствуют развитию
описанных выше факторов, определяющих общую
выносливость спортсмена?
Развитию дыхательной и кровеносной систем организма,
увеличению возможностей данных систем по доставке
кислорода к мышцам должны способствовать тренировки,
сопровождающиеся созданием максимальной потребности
мышц в кислороде. Такого рода нагрузка вызывает
напряжение указанных систем организма и,
соответственно, способствует необходимым
адаптационным изменениям в данных системах.
Высокая скорость потребления кислорода достигается
при нагрузках такой мощности, поддерживать которую
организм спортсмена способен лишь ограниченное
время, после чего наступает усталость. Поэтому
эффективными в данном случае будут серии
высокоинтенсивных нагрузок, перемежающиеся с
отдыхом, необходимым для восстановления сил. Время
удержания максимума потребления кислорода составляет
обычно не более шести минут — именно столько и
должно длиться тренирующее упражнение аэробной
направленности. Отдых между повторениями упражнения
в этом случае должен также составлять шесть минут.
Эффективными при воздействии на аэробные способности
организма оказываются и серии более коротких
высокоинтенсивных нагрузок длительностью от 30
до 90 секунд, чередующихся со столь же короткими
интервалами отдыха. Данный метод получил название
"циркуляторной интервальной тренировки", так как он
наиболее эффективно воздействует на циркуляторные
показатели кровеносной системы и вызывает выраженную
гипертрофию сердца. Эффективность метода заключается
в том, что потребление кислорода в первые минуты
отдыха после прекращения нагрузки сохраняется на
высоком уровне, поскольку происходит так называемый
возврат кислородного долга (получение окислительным
путём энергии, необходимой для восполнения
запасов АТФ и креатинфосфата, а также для вывода
молочной кислоты из мышц). Таким образом, в период
короткого отдыха уровень потребления кислорода
снижается несущественно, в то время как мышцы
восстанавливают силы, восполняя запасы АТФ и
креатинфосфата и избавляясь от продуктов метаболизма,
после чего получают возможность вновь развивать
высокое усилие и вновь создавать высокую потребность
в кислороде. Поэтому в течение всей циркуляторной
интервальной тренировки уровень потребления кислорода
совершает незначительные колебания возле максимальных
значений.
Для развития способности организма поддерживать
кислотно-щелочное равновесие крови (за счёт
ускорения утилизации кислых продуктов метаболизма
и за счёт накопления резервов буферных веществ)
необходимо в ходе тренировки добиваться
максимального повышения кислотности крови
(естественно, в пределах физиологически нормальных
величин). Для чего наиболее эффективны серии
высокоинтенсивных нагрузок длительностью 1-2 минуты
с 1-2 минутным интервалом отдыха между подходами.
Объясняется это тем, что максимум накопления
молочной кислоты в крови наблюдается через
некоторое время после прекращения короткой
высокоинтенсивной нагрузки. Задержка в достижении
максимума кислотности крови связана с необходимостью
какого-то времени на вывод молочной кислоты из
мышцы. Повторные нагрузки после отдыха,
достаточного для значительного вывода молочной
кислоты из мышц и восстановления их
работоспособности, но не столь длительного, чтобы
уровень кислоты в крови успел снизиться, приводят
к наложению максимумов выброса кислоты в кровь
друг на друга и к значительному сдвигу
кислотно-щелочного равновесия крови в кислую
сторону. Усталость мышц ввиду остаточного накопления
в них продуктов метаболизма наблюдается после
трёх-четырёх повторений такой нагрузки, поэтому
эффективно будет разделить тренировку на несколько
серий по 3-4 подхода с 10-15 минутным
отдыхом между сериями.
Теперь разберёмся с обеспечением мышц топливом.
Основными источниками энергии для мышечной
деятельности являются жирные кислоты, углеводы
(в основном, глюкоза) и аминокислоты. Запас
свободных аминокислот в организме весьма
незначителен, к использованию собственных белков
в качестве топлива организм прибегает только в
условиях недостатка энергии — например, в
условиях голодания или длительных истощающих
нагрузок. При этом аминокислоты, получаемые при
катаболизме собственных белков, всё равно, как
правило, проходят этап преобразования в печени в
глюкозу.
Таким образом, основными источниками энергии для
мышечной деятельности остаются жирные кислоты и
глюкоза. Жирные кислоты запасаются в
жировой ткани, а при необходимости извлекаются в
кровь и доставляются к работающим мышцам.
Саркоплазма мышц располагает и собственным
небольшим запасом жирных кислот. Запасы жиров в
организме практически неисчерпаемы в рамках
единичной тренировки, и если марафонский бег
обеспечивался бы исключительно жирными кислотами,
то для преодоления дистанции потребовалось бы
всего лишь около 320 граммов жира —
в то время как даже весьма худой человек располагает
несколькими килограммами жиров, а у отдельных
людей вес жировой ткани может достигать
нескольких десятков килограммов. К сожалению,
возможности жиров как источника энергии ограничены.
Жирные кислоты активно используются только при
низкоинтенсивных нагрузках, поскольку выход энергии
на одну молекулу кислорода и скорость окисления для
жиров несколько ниже, чем для глюкозы. Поэтому при
повышении энергозатрат митохондрии переключаются с
жирных кислот на глюкозу.
Более того, энергозатраты, превышающие окислительные
возможности мышц, активизируют гликолиз, и в этом
случае глюкоза становится незаменимым источником
энергии. Глюкоза запасается организмом главным
образом в мышцах в виде гранул гликогена. Какой-то
запас гликогена имеется и в печени —
100-200 граммов. При коротких интенсивных нагрузках
энергозатраты мышц покрываются за счёт внутренних
резервов гликогена. Размер внешних запасов энергии
становятся актуальным лишь при пролонгированных
нагрузках. Запасы жиров, как я уже отмечал ранее,
исчерпать нереально при любой разумной
длительности нагрузки, поэтому при использовании
жиров в качестве источника энергии имеет значение
не их количество, а активность ферментов,
извлекающих жирные кислоты из жировой ткани, и
скорость проникновения жирных кислот в митохондрии.
А вот резерв гликогена в печени может сыграть
решающее значение при длительных нагрузках, поэтому
только запасы гликогена, но не запасы жиров, и
можно рассматривать в качестве фактора,
ограничивающего общую выносливость организма.
Соответствующие тренировки способны привести к
увеличению запасов гликогена в печени и мышцах.
Происходит это увеличение по уже известной схеме:
истощение — восстановление —
сверхвосстановление. После истощающих нагрузок
и при условии достаточного потребления углеводов с
пищей, суперкомпенсация гликогена в печени и
мышцах наступает примерно на третьи сутки. Для
повышения содержания гликогена в печени
используется также метод "углеводной
загрузки", когда в течение нескольких дней
ограничивается потребление углеводов, а затем за
день до соревнований потребление углеводов
значительно увеличивают, что приводит к резкому
увеличению запасов гликогена в печени.
На этом я, пожалуй, завершу рассмотрение
тренировочных методов, воздействующих на факторы
общей выносливости организма, и перейду к
рассмотрению собственно силовой выносливости мышц.
Способность мышц сокращаться с требуемым усилием
определяется прежде всего насыщенностью мышц
энергией. И хотя основной причиной снижения силы
сокращения мышц является вовсе не отсутствие АТФ,
а снижение АТФазной активности миозина и
нарушения в механизме передачи возбуждения с
нерва вглубь волокна, причиной упомянутых
нарушений являются метаболические факторы
утомления (молочная кислота, ортофосфорная
кислота, АДФ и др.), а
их появление в мышце связано как раз с доступностью
энергии. Недостаток АТФ, производимой
окислительным путём, приводит к активизации
гликолиза и к появлению в мышце большого количества
молочной кислоты (лактата). Недостаток энергии,
производимой путём гликолиза, приводит к
истощению запасов креатинфосфата и, соответственно,
к увеличению в мышце концентрации ортофосфата.
По Н.И.Волкову, при рассмотрении факторов
работоспособности мышц в зависимости от основного
механизма энергообеспечения следует различать
аэробную (окисление) и анаэробную
работоспособности. Анаэробная работоспособность,
в свою очередь, делится на лактатную (гликолиз)
и алактатную (креатинфосфат). В качестве главных
критериев оценки механизмов энергообеспечения
мышечной деятельности принято выделять
максимальную мощность, время удержания
максимальной мощности и общую ёмкость механизма.
Максимальная мощность — это наибольшая скорость
образования АТФ в данном метаболическом процессе.
От мощности механизма энергообеспечения зависит
возможная сила сокращения мышц в данном режиме
работы. Под ёмкостью понимается общее количество
энергии, которое можно получить за счёт данного
механизма ресинтеза АТФ.
Алактатная работоспособность мышц
Максимальная алактатная мощность, с одной
стороны, зависит от концентрации и активности
фермента креатинкиназы (переносящего фосфатную
группу с креатинфосфата на АДФ) и собственно
креатинфосфата. С другой стороны, мощность данной
реакции зависит от потребности мышц в энергии и,
соответственно, определяется максимальной
скоростью расходования АТФ, развиваемой мышцами.
Максимальная длительность удержания алактатной
мощности составляет 6-12 секунд. Алактатная
ёмкость зависит от запасов креатинфосфата в мышце.
О методах тренировки алактатной мощности и ёмкости
я уже рассказывал ранее, рассматривая методы
развития максимальной силы, и сейчас не буду
подробно останавливаться на этом вопросе.
Лактатная работоспособность мышц
Максимальная лактатная мощность определяется
главным образом концентрацией и активностью
ключевых ферментов гликолиза. Время удержания
максимальной мощности данного метаболического
процесса составляет 30-60 секунд и определяется,
с одной стороны, устойчивостью ферментов гликолиза
к понижению рН среды (повышение кислотности среды
ингибирует активность гликолитических ферментов,
что подавляет энергопроизводство), и устойчивостью
кислотно-щелочного равновесия внутренней среды
мышц в условиях усиленной выработки лактата. С
другой стороны, время удержания максимальной
гликолитической мощности лимитируется факторами
утомления мышцы, снижающими интенсивность
сокращения.
Из написанного выше следует, что для запуска
адаптационных процессов, направленных на
увеличение максимальной гликолитической мощности,
длительность нагрузки должна соответствовать
времени удержания максимальной мощности данного
метаболического процесса, что составляет
30-60 секунд. Отдых между подходами должен быть
достаточно длительным для обеспечения вывода
продуктов метаболизма из мышцы и для развития
высокой мощности гликолиза в следующем подходе.
Устойчивость рН среды мышечных волокон
к выбросу молочной кислоты и устойчивость ключевых
ферментов к снижению рН вырабатывается в ходе
тренировок, сопровождающихся максимальным
накоплением лактата в мышцах. Это могут быть
нагрузки высокой интенсивности, длительностью
1-1,5 минуты до наступления отказа мышц,
вызванного сильным закислением, либо более
короткие нагрузки длительностью 20-40 секунд
со столь же коротким интервалом отдыха, приводящие
к кумулятивному накоплению лактата в мышцах.
Гликолитическая ёмкость определяется главным
образом запасами гликогена в мышцах, гликоген
печени для процессов гликолиза не обладает
достаточной мобильностью. О методах накопления
мышечного гликогена, равно как и гликогена
печени, я уже рассказывал при рассмотрении
факторов общей работоспособности организма.
Аэробная работоспособность мышц
Максимальная аэробная мощность зависит главным
образом от плотности митохондрий в мышечных
волокнах, от концентрации и активности
окислительных ферментов, от скорости поступления
кислорода вглубь волокна. Объём кислорода,
доступного для окислительных реакций,
лимитируется как факторами общей
работоспособности организма, которые я ранее уже
рассматривал, так и рядом локальных
внутримышечных факторов. Среди них можно
выделить капилляризацию мышц, концентрацию
миоглобина, диаметр мышечного волокна (чем меньше
диаметр волокна, тем лучше оно снабжается
кислородом и тем выше его относительная аэробная
мощность). Скорость производства АТФ за счёт
окисления достигает максимальных значений
на второй-третьей минутах работы, что связано с
необходимостью развёртывания множества процессов,
обеспечивающих доставку кислорода к митохондриям.
Время удержания максимальной аэробной мощности
составляет примерно 6 минут, в дальнейшем
аэробная мощность снижается по причине усталости
всех активно работающих систем организма.
Соответственно, для повышения аэробной мощности
мышц тренировочная нагрузка должна длиться не
менее двух минут (для выхода скорости
энергопроизводства на максимум). Не имеет смысла
и затягивать нагрузку дольше, чем на 6 минут,
при тренировке именно мощности, поскольку далее
происходит её (мощности) снижение. Эффективным
оказывается многократное повторение таких
нагрузок.
В заключение приведу сводную таблицу
тренировочного воздействия на работоспособность
мышц в различных режимах работы, почерпнутую
мной из диссертации М.Хосни, посвящённой изучению
биохимических основ интервальной тренировки. Для
развития соответствующих качеств Хосни
рекомендует следующие методические приёмы:
| Направление воздействия тренировки |
Интенсивность |
Длительность нагрузки |
Отдых между подходами |
Количество подходов |
| Алактатная анаэробная мощность |
Максимальная |
7-10 сек. |
2-5 мин. |
5-6 |
| Алактатная анаэробная ёмкость |
Максимальная |
7-10 сек. |
0,3-1,5 мин. |
10-12 |
| Лактатная анаэробная мощность |
Высокая |
20-30 сек. |
6-10 мин. |
3-4 |
| Лактатная анаэробная ёмкость |
Высокая |
40-90 сек. |
5-6 мин. |
10-15 |
| Аэробная мощность |
На максимуме потребления кислорода |
0,5-2,5 мин. |
0,5-3 мин. |
10-15 |
| Аэробная ёмкость |
На максимуме потребления кислорода |
1-6 мин. |
1-6 мин. |
Больше 10 |
На этом я заканчиваю изложение основ тренировки
работоспособности мышц и перехожу к анализу
основных факторов, определяющих мышечные объёмы
спортсмена.
Часть VI. Рассмотрение
тренировочных методик
Ну что же, основные методы тренировок,
способствующих развитию силы и силовой
выносливости мышц, мной уже рассмотрены. Настало
время приступить к рассмотрению тренировочных
методик, в полной мере способствующих гипертрофии
мышц, для чего следует определить тканевые и
внутриклеточные структуры, от развития которых
зависят мышечные объёмы спортсмена. Я немного уже
затрагивал этот вопрос во второй части, теперь
остановлюсь на нём подробнее.
Как отмечалось, объём мышцы определяется прежде
всего количеством мышечных волокон (клеток) в
теле мышцы, размером самих этих волокон, а также
объёмом межклеточного вещества, представленного
главным образом кровеносными сосудами и
соединительной тканью, отделяющей друг от друга
отдельные волокна и их пучки. Немаловажное
значение для визуальных объёмов спортсмена
имеют и запасы жира в организме, однако вклад
жиров уже трудно назвать вкладом именно в
"мышечные" объёмы, а стандарты соревновательного
бодибилдинга требует минимизации такого "вклада"
— поэтому методы тренировок, приводящих к
увеличению жировой составляющей объёмов
спортсмена, я рассматривать не буду: они и так
хорошо всем известны. Увеличение количества
мышечных волокон у человека ни разу достоверно
не фиксировалось в экспериментах, хотя, как я уже
отмечал выше, гиперплазия не кажется мне столь уж
невероятным явлением после того, как она была
зафиксирована у подопытных животных. Но, дабы не
прослыть пустым фантазёром, я не хочу включать
гиперплазию в число факторов мышечной гипертрофии
до появления достоверных экспериментов,
зафиксировавших увеличение количества мышечных
волокон у человека.
Итак, нам, людям, остаётся уповать только на
капилляризацию мышц, на увеличение в объёме
мышечных волокон и на рост соединительной ткани.
Объём мышечных волокон контролируется прежде
всего количеством мышечных ядер в волокне.
Именно от количества ядер при прочих равных
условиях зависит общий объём белка, синтезируемого
мышечным волокном в единицу времени. И этот
фактор незаслуженно игнорируется многими
специалистами при рассмотрении причин мышечной
гипертрофии под действием тренировок.
Как отмечалось, к увеличению количества мышечных
ядер приводит деление клеток-сателлитов,
инициируемое факторами, появляющимися в мышечном
волокне при его повреждении. Но ядра служат лишь
первопричиной, вообще же увеличивают объём
волокна совсем иные клеточные структуры —
такие, как миофибриллы, саркоплазма, митохондрии
и др. Вот какие данные о потенциале роста мышц за
счёт различных клеточных и межклеточных структур
привёл Фред Хэтфилд:
Примерный вклад различных
факторов в увеличение размеров мышцы, %:
- Капилляризация — 3-5
- Митохондрии — 15-25
- Саркоплазма (клеточная жидкость) — 20-30
- Соединительные ткани — 2-3
- Мышечные фибриллы — 20-30
- Гликоген — 2-5
Как можно видеть, существенный вклад в объём мышц
вносит количество и поперечное сечение миофибрилл
в мышечном волокне. Сравнимое влияние на размеры
мышц оказывает объём саркоплазмы и расположенных
в ней митохондрий. Таким образом, следует
различать миофибриллярную и саркоплазматическую
гипертрофию. На первый взгляд, потенциал
саркоплазматической гипертрофии (клеточная
жидкость + митохондрии + гликоген) даже превышает
потенциал роста за счёт сократительных структур.
Но при внимательном рассмотрении становится ясно,
что саркоплазматическая гипертрофия находится в
подчинённом отношении к миофибриллярной.
Каждая миофибрилла требует наличия в клетке
определённого объёма саркоплазмы и митохондрий,
призванных обеспечивать их (миофибрилл)
функционирование. Рост миофибриллярных структур
автоматически приводит к соответствующему
увеличению саркоплазматических структур. Более
того, процентные отношения, приведённые Хэтфилдом,
вызывают некоторые сомнения — тем более что
Хэтфилд не указал источник своей информации. Так,
в учебнике биологической химии за авторством
Т.Т.Берёзова и Б.Ф.Коровкина приведены несколько
иные сведения. Химический анализ мышечной ткани
показывает, что 70-80% мышечной массы приходится на
воду, а 20-30% — это сухой остаток, состоящий из
белков, липидов и углеводов. Процентное соотношение
белков, обнаруживаемых в сухом остатке, следующее:
сократительные белки — 35%, белки саркоплазмы
— 45%, белки стромы (соединительной ткани)
— 20%. То есть
процентные соотношения белков получаются близкими к
приведённым Хэтфилдом, однако не следует забывать,
что это отношения массы, а не объёма. По данным
этого же источника, миофибриллы занимают около 80%
объёма мышечного волокна, то есть на все остальные
структуры помимо собственно миофибрилл суммарно
приходится не более 20% объёма клетки. Таким
образом, соотношение между миофибриллярной
и саркоплазматической гипертрофиями получается уже
несколько иным, чем это следует из данных,
приведённых Хэтфилдом: миофибриллярная гипертрофия
может дать до 80% прироста объёмов волокна, а
саркоплазматическая гипертрофия — только 20%.
Но человеку, стремящемуся к максимальному развитию
мускулатуры, не следует пренебрегать и этими
двадцатью процентами.
Понятно, что относительный объём саркоплазмы
мышечной клетки зависит и от активности
использования миофибрилл, то есть от объёма регулярно
производимой мышцами работы. Связь между
концентрацией митохондрий в клетке с её
энергетическими потребностями, я думаю, не вызывает
вопросов, а вот почему увеличение энергозатрат
увеличивает объём саркоплазмы мышечной клетки,
стоит разъяснить. Саркоплазма — это не только
клеточная жидкость (вода), это ещё и миллионы
миллионов молекул различных веществ, взвешенных и
растворённых в ней. Это, прежде всего, крупные
молекулы белков-ферментов, призванные обеспечивать
протекание множества жизненно важных химических
реакций, в том числе и энергообеспечивающих. Это
запасы органического топлива — АТФ,
креатинфосфата, гликогена, жирных кислот и
аминокислот. Это молекулы миоглобина. Это, наконец,
всевозможные ионы (К+, Са++,
Na+, Mg++ и др.).
Но основной объём саркоплазмы создают даже не сами
перечисленные вещества, а окружающая их вода.
Вещества, растворённые и взвешенные в саркоплазме,
одним своим наличием связывают и задерживают в
клетке определённое количество молекул воды.
Накопление в клетке перечисленных выше веществ
пропорционально увеличивает объём саркоплазмы. С
эффектом резкого увеличения объёма саркоплазмы
мышечных волокон во время тренировки за счёт
жидкости, нахлынувшей в клетки из межклеточного
пространства и плазмы крови, хорошо знакомы все
занимающиеся. При гликолизе, активизирующемся во
время мышечной деятельности, глюкоза распадается
на молочную кислоту в соотношении 1:2 (одна
молекула глюкозы — две молекулы молочной
кислоты). Поскольку две молекулы кислоты связывают
больше молекул воды, чем одна молекула глюкозы,
то активизация гликолиза увеличивает потребность
клетки в жидкости, и вода устремляется внутрь
мышечных волокон, что приводит к их разбуханию
и заметному увеличению мышцы в объёме. Однако не
следует путать такое временное увеличение объёма
с мышечным ростом, ибо едва только молочная
кислота выводится из мышцы, объём клеточной
жидкости сразу возвращается к норме. Гипертрофией
саркоплазмы можно считать лишь устойчивое
увеличение её объёма, фиксируемое в состоянии
покоя.
Увеличение объёма саркоплазмы может происходить
не только благодаря простому накоплению в ней
перечисленных выше веществ. Саркоплазма мышечных
волокон несколько отличается от саркоплазмы иных
клеток, и связано это с наличием в мышечных
волокнах таких структур, как миофибриллы. Каждая
миофибрилла окружена плотной сетью
саркоплазматического ретикулума, состоящей из
терминальных цистерн с ионами Ca++ (ионы
выбрасываются в саркоплазму при сокращении), и
переплетения так называемых Т-трубочек,
связывающих терминальные цистерны с сарколеммой
(оболочкой волокна) и обеспечивающих поступление
сигнала к сокращению. То есть каждая миофибрилла
окружена строго определённым объёмом
саркоплазматических структур. Объём этих структур
пропорционален площади поверхности миофибрилл в
волокне. Соответственно, чем больше диаметр
отдельных миофибрилл, тем меньше объём окружающей
миофибриллу саркоплазмы по отношению к объёму
сократительных белков внутри этой миофибриллы
(выше доля сократительных белков в волокне). Но
чем больше объём каждой миофибриллы, тем труднее
обеспечивать её энергопотребности, так как
длиннее путь переноса энергии от поверхности
миофибриллы (где расположены основные источники
энергии — митохондрии) вглубь. Соответственно,
при активации мышц адаптация волокон к изменению
условий жизнедеятельности может быть направлена
на расщепление крупных миофибрилл на несколько
мелких. В случае расщепления миофибрилл их масса
остаётся неизменной, однако возрастает их
количество и, соответственно, увеличивается
площадь поверхности миофибрилл, что неминуемо
должно сопровождаться увеличением объёма
саркоплазматического ретикулума. То есть в этом
случае происходит гипертрофия мышечной клетки без
увеличения объёма сократительных белков —
иными словами, наблюдается саркоплазматическая
гипертрофия. Имея в виду то, что объём
саркоплазмы мышечного волокна может быть увеличен
как благодаря накоплению различных веществ,
ответственных за энергопроизводство клетки, так и
благодаря расщеплению миофибрилл в процессе
эргономического приспособления к увеличивающемуся
объёму работы, можно утверждать, что
саркоплазматическая гипертрофия является
адаптационной реакцией мышц на увеличение объёма
работы, регулярно выполняемого мышцами.
Из вышеприведённого краткого анализа становится
ясно, что никаких особых специфических методов
тренировки, направленных исключительно на
увеличение объёмов мышц, не существует.
Гипертрофии мышц в той или иной мере способствуют
все рассмотренные ранее тренировочные методики,
направленные на развитие силы (за счёт развития
сократительных структур) и силовой выносливости
мышц (саркоплазматическая гипертрофия). Точнее,
развитие ряда клеточных структур может вносить
вклад в развитие таких мышечных качеств, как
сила, силовая выносливость и объём (см. рис. 1).
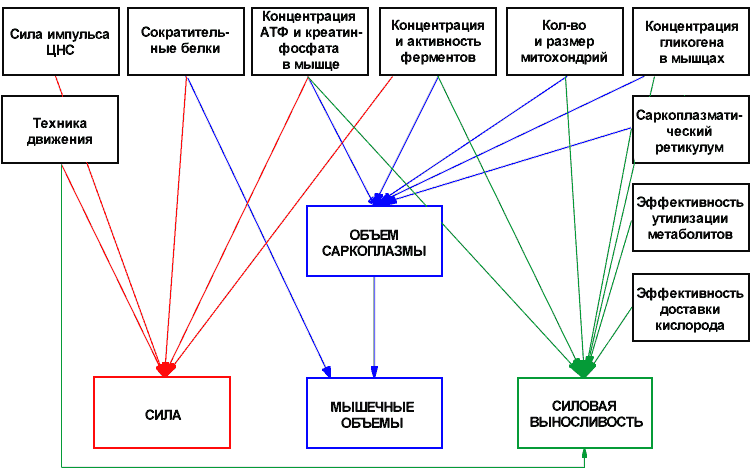 Рисунок 1
Для максимального увеличения силы, выносливости и
объёма мышц следует использовать тренировки,
воздействующие на все основные факторы, вносящие
вклад в развитие упомянутых качеств.
Взаимодействие срочных тренировочных эффектов
Каким образом можно объединить различные
тренировочные методики в общий тренировочный
процесс? Основная проблема состоит здесь в том,
что далеко не все методические приёмы совместимы
в ходе одного тренировочного занятия.
Адаптационные изменения в организме под влиянием
нагрузок разной направленности могут быть как
усилены, так и ослаблены. Этот феномен принято
называть взаимодействием срочных тренировочных
эффектов. Взаимодействие тренировочных эффектов
разнонаправленных нагрузок может быть
положительным, отрицательным и нейтральным.
Наиболее выраженные адаптационные изменения в
организме формируются при положительном
взаимодействии тренировочных эффектов. При
неправильном построении тренировок отрицательное
взаимодействие нагрузок существенно снижает
общий тренировочный эффект и даже может
привести к перетренированности. По данным
исследований, проведённых Н.И.Волковым,
положительный эффект от взаимодействия
разнонаправленных нагрузок может быть достигнут
только в ограниченном числе их сочетаний
(см. таблицу).
| Последовательность выполнения нагрузок |
Характер достигаемого срочного тренировочного эффекта |
| Алактатного анаэробного + гликолитического анаэробного воздействия | Гликолитический анаэробный |
| Алактатного анаэробного + аэробного воздействия | Аэробный |
| Гликолитического анаэробного (в небольшом объёме) + аэробного воздействия |
Аэробный |
| Аэробного (в небольшом объёме) + алактатного анаэробного воздействия | Алактатный анаэробный |
Взаимодействие отставленных тренировочных эффектов
Следующая важная проблема, возникающая при
построении тренировочного процесса, кроется в
различном времени, требующемся для восстановления
нагружаемых в ходе тренировки функциональных
систем организма. Хорошо известно, что быстрее
всего после тренировки восстанавливаются
алактатные резервы организма, чуть дольше
происходит восстановление функциональных систем,
ответственных за мощность и ёмкость
гликолитических процессов (до трёх суток), а
больше всего времени требуется на восстановление
разрушенных в ходе тренировки клеточных структур.
Данный феномен получил название гетерохронизма
восстановительных процессов. Здесь я рекомендую
обратиться ко второй части моего текста и
вспомнить последовательность прохождения
восстановительных процессов в организме.
Снижение тренируемой функции под действием
тренировки => восстановление (компенсация) =>
сверхвосстановление (суперкомпенсация) =>
утраченное сверхвосстановление (декомпенсация).
Положительное взаимодействие тренировочных
эффектов наблюдается лишь при задании новой
тренировочной нагрузки в состоянии
сверхвосстановления (при возросших функциональных
возможностях). Слишком большой перерыв между
тренировками приводит к воздействию на
тренируемую функцию в состоянии утраченного
сверхвосстановления и не способствует закреплению
адаптационных изменений, вызванных предыдущими
тренировками. Недостаточный отдых между
тренировками приводит к тому, что нагрузка на
тренируемую функцию задаётся ещё до того, как
функция восстановится после предыдущей
тренировки, что при длительном повторении такой
ситуации может стать причиной перетренированности.
Поэтому тренировочный процесс строится по
возможности таким образом, чтобы в период
восстановления одной тренируемой функции
задаваемая нагрузка воздействовала на иную
систему организма и не оказывала отрицательного
воздействия на восстанавливаемую функцию.
Например, после объёмной тренировки аэробной
направленности восстановление энергетического
потенциала мышц может занять около трёх дней, в
это время вполне возможно проведение небольших
по объёму тренировочных занятий анаэробного
характера. Но тренировки далеко не всегда можно
разделить столь чётко по направленности
воздействия. При одновременном напряжении
различных функциональных систем, не обладающих
синхронностью восстановительных процессов,
указать оптимальный перерыв между тренировочными
нагрузками становится невозможно. Какое бы время
отдыха атлет ни выбрал, оно будет оптимальным
лишь для одних функциональных систем, а другие
функциональные системы обязательно окажутся либо
в состоянии недовосстановления, либо в состоянии
утраченного сверхвосстановления. В той или иной
мере любая тренировка воздействует на системы,
ответственные и за алактатные, и за
гликолитические, и за аэробные процессы в мышцах
и организме. Более того, достаточно интенсивная
тренировка в анаэробном режиме оказывает, как
правило, разрушающее воздействие на сократительные
структуры мышечных волокон, а интенсивная аэробная
тренировка может оказать аналогичное разрушающее
воздействие на митохондрии мышечных клеток.
Восстановление клеточных структур требует больше
времени, чем необходимо для достижения
сверхвосстановления энергетических компонентов; в
свою очередь, различные энергетические компоненты
также имеют различные сроки достижения состояния
сверхвосстановления. Как же тут быть?
О самом простом решении данной проблемы я уже
писал: достаточно проводить тренировки с частотой,
обеспечивающей достижение состояния
суперкомпенсации самой долговосстанавливающейся
функциональной системы. Такие тренировки обеспечат
планомерный рост данной функции и позволят
гарантировано избежать состояния
перетренированности, поскольку ни одна
функциональная система не будет нагружаться в
состоянии недовосстановления. В то же время ряд
функциональных систем к моменту новой тренировки
окажется в состоянии декомпенсации, что не
способно привести к развитию долговременной
адаптации данных систем (см. рис. 2).
Как можно понять, я веду речь о тренировках в
стиле "Супертренинга" Майка Менцера. Данный метод
подходит только для развития силы и массы мышц за
счёт роста сократительных структур клетки (именно
восстановление и суперкомпенсация сократительных
белков клетки, разрушенных в ходе тренировки,
требует наибольшего времени). Но хотя развитие
миофибриллярного аппарата мышечных волокон
является определяющим для силы и массы мышц,
добиться максимально возможного развития силы,
существенной саркоплазматической гипертрофии,
значительного повышения силовой выносливости и
общей работоспособности организма таким методом
тренировки невозможно.
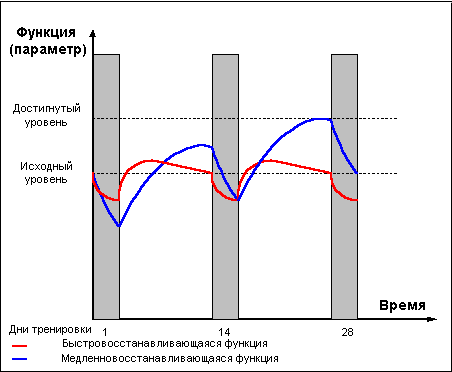 Рисунок 2
Совместить миофибрильную гипертрофию с повышением
работоспособности мышц и тем самым увеличить долю
саркоплазматической гипертрофии можно посредством
метода регулирования интенсивности нагрузки.
Наибольшим воздействием на сократительные
структуры мышечных волокон, как отмечалось,
обладают высокоинтенсивные тренировки, в ходе
которых мышцы развивают относительно высокую
скорость расходования энергии. Тренировки, обладающие
таким разрушающим мышечные структуры эффектом и
запускающие соответствующие адаптационные
процессы, следует проводить только тогда, когда
мышцы войдут в состояние сверхвосстановления, то
есть после полного восстановления разрушенных
интенсивной тренировкой клеточных структур. Как
показывает опыт, в зависимости от нагрузки и
индивидуальных восстановительных особенностей
спортсмена для полного восстановления и достижения
состояния суперкомпенсации мышцам может
потребоваться до двух-трёх недель. Но в течение
этого периода для обеспечения полноценного
восстановления вовсе не обязательно абсолютно
исключать нагрузку на мышцы. Для того чтобы мышцы
не вошли в состояние декомпенсации энергетических
функций, период отдыха между тяжёлыми тренировками
можно заполнить тренировками меньшей интенсивности,
не оказывающими на мышцы разрушающего воздействия
(либо сводящими такое воздействие к минимуму) и в
то же время способствующих закреплению
адаптационных изменений в энергообеспечивающей
системе мышц и организма в целом. В этом случае
тренировочный процесс разбивается на мезоциклы,
состоящие из двух микроциклов — ударного и
восстановительного (см. рис. 3).
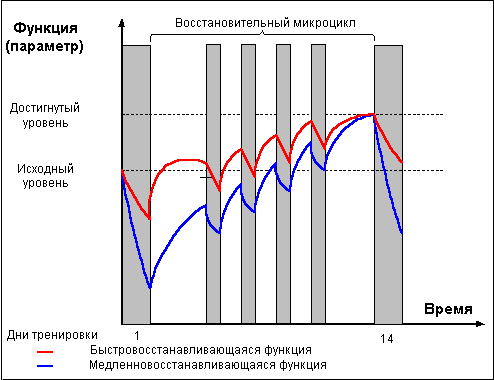 Рисунок 3
Ударный микроцикл может состоять из нескольких
высокоинтенсивных занятий, в ходе которых
задаётся нагрузка на все основные мышечные
группы (обычно длительность микроцикла составляет
одну неделю). За ударным микроциклом следует
восстановительный, длительностью около двух
недель (возможны отступления в бОльшую либо
меньшую стороны в зависимости от восстановительных
возможностей индивида и интенсивности ударного
микроцикла). Во время восстановительного
микроцикла нагрузка на одну и ту же мышцу
задаётся не реже, чем один раз в неделю и не
чаще, чем раз в 2-3 дня. При этом по сравнению с
ударным микроциклом снижается либо средняя
интенсивность тренировки, либо объём работы, либо
оба параметра одновременно.
Схожие принципы прослеживаются и в рекомендациях
Ф.Хэтфилда применительно к тренировкам
бодибилдеров. Хэтфилд предложил чередовать для
каждой мышечной группы тяжёлые, средние и лёгкие
тренировки в такой последовательности: тяжёлая
тренировка — средняя тренировка — лёгкая
тренировка — средняя тренировка — тяжёлая
тренировка. Отдых после тяжёлой тренировки должен
составлять 4-5 дней, после средней
— 3-4 дня, и 2-3 дня
после лёгкой тренировки. Длительность отдыха,
по мнению Хэтфилда, может немного колебаться
в зависимости от тренируемой мышечной
группы (например, ноги — опять же по мнению
Хэтфилда — требуют более длительного отдыха,
чем руки). Хотя Хэтфилд чётко не разделил
тренировочный процесс по микроциклам, а предложил
проводить тренировки различных мышечных групп
независимо друг от друга, получая во время одного
тренировочного занятия сложную комбинацию из
лёгких, средних и тяжёлых тренировок для
различных мышц, в его схеме всё же
прослеживается микроциклирование нагрузки —
если рассматривать тренировки одной мышечной
группы (см. рис. 4).
Несмотря на собственные рекомендации о
целесообразности применения средних и лёгких
тренировок, Хэтфилд явно полагает, что за 4-5 дней
после тяжёлых тренировок происходит полное
восстановление мышц (впрочем, возможно, всё дело в
неудачной формулировке или переводе текстов
Хэтфилда). Будь это так, необходимости в средних и
лёгких тренировках не возникло бы, а после
четвёртого-пятого дня можно было бы сразу
проводить новую тяжёлую тренировку. На самом же
деле, как я уже не раз отмечал, для полного
восстановления мышц после интенсивных нагрузок
требуется гораздо больше времени. И если
просуммировать дни отдыха между всеми типами
тренировок в системе Хэтфилда, то получится, что
полное время восстановления между тяжёлыми
тренировками составит около двух недель. Именно
столько времени отводится на отдых между
тренировками одной мышцы в "Супертренинге" Майка
Менцера.
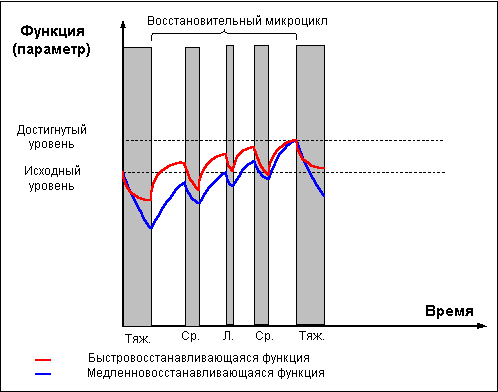 Рисунок 4
Часть VII. Принципы
чередования ударных и восстановительных
микроциклов
До сих пор я рассматривал тренировочные методики,
обеспечивающие мышечную гипертрофию путём
последовательного достижения сверхвосстановления
мышц и организма в целом к моменту каждой
напряжённой (пиковой) тренировки. Но мышечная
гипертрофия возможна и в тех случаях, когда
напряжённые тренировки проводятся задолго до
полного восстановления мышц после предшествующей
нагрузки. Проведение нескольких напряжённых
тренировок подряд, приводящих к
последовательному снижению функциональных
возможностей спортсмена, вполне допустимо, если
за серией ударных нагрузок следует
восстановительный микроцикл (серия тренировок
меньшей напряжённости), что позволяет достичь
суперкомпенсации тренируемых функций и выйти на
новый уровень функциональных возможностей
(см. рис. 5).
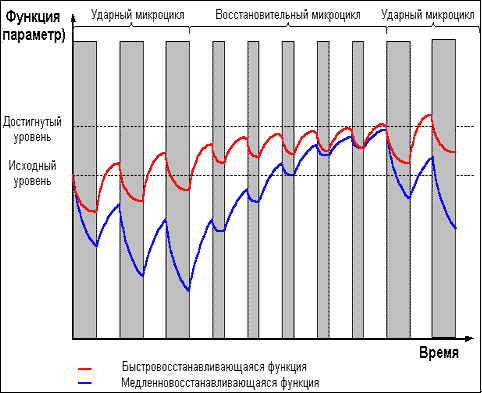 Рисунок 5
На принципе чередования ударных и
восстановительных микроциклов основано
большинство тренировочных методик "классической"
спортивной школы, характеризующихся, как
правило, очень высокой частотой тренировочных
занятий и большим объёмом тренировочных нагрузок,
что (частота тренировок и объём нагрузок)
объясняется зависимостью результатов в
большинстве видов спорта от уровня общей и
специфической работоспособности спортсмена.
Применительно к силовым видам спорта критерием
оценки напряжённости тренировочного занятия при
таком подходе служит тоннаж тренировки I = i x n,
где n — количество поднятых штанг, а
i — средний вес штанги или так называемая
"средняя интенсивность за тренировку",
измеряемая в килограммах.
Примером системы тренировок бодибилдеров,
основанной на принципах классической теории
физической культуры, может служить методика
Владимира Гончарова, изложенная им в книге
"Логика тренинга". Она, в отличие от большинства
"объёмных" методик, публикуемых в популярных
журналах, единственным принципом которых
является принцип "бери больше, кидай дальше",
действительно имеет внутреннюю логику и
заслуживает внимания. Каждая мышца, по методике
Гончарова, нагружается не реже двух раз в
неделю, а общее количество тренировочных
занятий в зависимости от уровня тренированности
может доходить до четырнадцати в неделю, то есть
до двух тренировок ежедневно. Естественно, ни о
каком восстановлении, а уж тем более
сверхвосстановлении мышц к следующему
тренировочному занятию при такой частоте
тренировок и речи быть не может. Ключевым
пунктом в методике Гончарова является так
называемый "функциональный спад" — состояние
снижения функциональных возможностей спортсмена,
в которое он неминуемо попадает по причине
накопления от тренировки к тренировке
остаточного недовосстановления. При появлении
признаков функционального спада (то есть при
невозможности выполнить запланированный объём
работы) производится снижение нагрузки (это
соответствует восстановительному микроциклу на
рисунке 5), благодаря чему происходит
восстановление и сверхвосстановление тренируемых
функций, что в дальнейшем позволяет спортсмену
закрепиться на новом объёме тренировочных
нагрузок, то есть выдерживать бОльшую нагрузку
без функционального спада.
Здесь следует напомнить, что дольше всего
длится восстановление сократительных белков и
иных структур мышечных клеток, разрушенных в
ходе тренировки, поэтому под медленно
восстанавливающейся функцией, обозначенной
синим цветом, здесь и на приведённых ранее
рисунках следует понимать совокупный объём
клеточных структур мышцы. Быстро
восстанавливающаяся функция на приведённых
рисунках — это обобщённый показатель энергетики
мышц (совокупный алактатный, гликолитический и
окислительный потенциалы). Как отмечалось во
второй части данного текста, адаптация мышц в
энергетическом плане приводит к снижению
разрушительного эффекта тренировки, именно
поэтому адаптация мышц к определённому объёму
работы приводит к тому, что данная нагрузка не
оказывает существенного разрушительного
воздействия на клеточные структуры мышц и,
соответственно, снижается потребность спортсмена
в отдыхе после тренировки — что даёт
возможность поддерживать достигнутый уровень
нагрузки без функционального спада. Для перехода
на новый уровень работоспособности мышц
увеличивается тоннаж при сохраняющейся частоте
тренировок, что вновь приводит к функциональному
спаду, и процесс повторяется. Итак, вот
схематичная последовательность развития
тренировочного процесса по методике Гончарова:
Увеличение нагрузки, приводящее к состоянию
функционального спада => снижение нагрузки
(восстановительный микроцикл) =>
сверхвосстановление => повышение нагрузки в
состоянии сверхвосстановления и закрепление на
новом уровне нагрузки без функционального спада.
Следует отметить, что эффект снижения
воздействия тренировки на целостность клеточных
структур по мере развития адаптационных
процессов в энергетике мышц позволяет достичь
состояния сверхвосстановления даже без снижения
уровня тренировочной нагрузки (без
восстановительного микроцикла). Например, когда
новичок, согласно устоявшимся традициям, сразу
начинает тренироваться 2-3 раза в неделю,
особенно не мудрствуя со временем восстановления
и циклированием нагрузки, он тем не менее
добивается значительного прогресса. Мышцы
новичка, конечно же, не успевают
восстанавливаться между тренировками
(подтверждением чему служит мышечная боль, не
прошедшая к моменту новой тренировки), а новичок
вновь и вновь нагружает мышцы через боль до тех
пор, пока энергетика мышц не адаптируются к
задаваемой нагрузке (сигналом чему служит
исчезновение или заметное снижение
посттренировочной боли), после чего и становится
возможным сверхвосстановление мышц. Повышение
нагрузки путём увеличения веса снаряда или
количества выполняемых за тренировку подъёмов
штанги усиливает воздействие тренировки на мышцы
и даёт новый стимул к сверхвосстановлению,
которое становится возможным без
восстановительного микроцикла только после
адаптации мышц к новому уровню нагрузки.
В связи с этим у кого-нибудь может возникнуть
следующий вопрос: зачем я морочил читателю голову,
рассказывая о различных тренируемых функциях, о
гетерохронизме восстановительных процессов, о
циклировании нагрузки и пр., если
сверхвосстановление мышц прекрасно происходит и
без всех этих хитрых вещей? Однако искушённый
читатель, конечно же, понимает, что прогресс при
тренировках такого рода возможен только в самом
начале спортивной карьеры — и практика это
неумолимо доказывает.
Во-первых, по мере увеличения размеров мускулатуры
возрастают и требования к системному обеспечению
восстановительных процессов (я имею в виду
гормональный фон, обеспеченность энергией и
аминокислотами и т.д.), эффективность восстановления
после микротравм постепенно снижается,
соответственно снижается и размер
суперкомпенсационной прибавки. Ясно, что постоянное
прерывание процесса восстановление мышц новыми
напряжёнными тренировками не сказывается
положительно на результате восстановления и может
стать причиной замедления роста результатов.
Во-вторых, очень скоро адаптация в энергетике мышц
достигает такого уровня, что тренировочные нагрузки
перестают вызывать микротравмы мышц, достаточные
для стимулирования восстановительных процессов
такой интенсивности, чтобы обеспечить значительное
сверхвосстановление. По этим причинам мышцы со
временем входят в состояние, называемое
"тренировочное плато" (рис. 6).
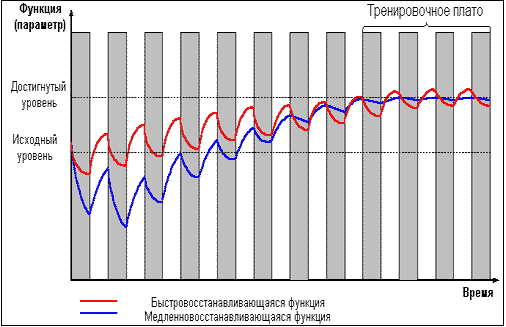 Рисунок 6
На этом рисунке для простоты изображён только один
цикл функционального спада и сверхвосстановления
перед входом в состояние "тренировочного плато"; на
практике спортсмен проходит, конечно, через ряд
таких циклов, прежде чем окончательно утрачивает
возможность дальнейшего прогресса от подобных
тренировок.
Итак, тренировка каждой мышцы не реже двух раз в
неделю с неизменно высокой интенсивностью и
стремлением к её (интенсивности) постоянному
увеличению, с одной стороны, не обеспечивает
максимально эффективного восстановления, а с
другой стороны, рано или поздно приводит к
излишней адаптированности мышц к нагрузке — к
состоянию "тренировочного плато". И поскольку
такой стиль тренировки является самым
распространённым по всем спортивным залам, то
получается парадоксальная ситуация: большинство
любителей "железного спорта" используют наименее
эффективные принципы построения тренировочного
процесса.
Если потребность в циклировании нагрузки и в
использовании восстановительных микроциклов при
высокой частоте тренировочных занятий хорошо
известна спортивным специалистам, и в неведении
остаётся лишь большинство любителей, то
упоминаний о влиянии адаптационных процессов в
энергетике мышц на чувствительность
сократительных структур к тренировке я не
встречал даже в специализированной литературе. К
моменту начала написания данного текста мне
казалось, что я единственный, кто пришёл к выводу
о необходимости дезадаптации мышц в энергетической
сфере для повышения эффективности воздействия
тренировки на рост мышечной массы.
Однако совсем недавно один мой друг, хорошо
знакомый с англоязычными интернет-сайтами
спортивной тематики, обратил моё внимание на
методику тренировки, разработанную Брайаном
Хейкоком (Bryan Haycock) и названную автором
HST (Hypertrophy Specific Training).
Значительную роль в тренировочном процессе
Хейкок отвёл периодической дезадаптации мышц, для
чего, с его точки зрения, нужно полностью
отказаться от тренировок в течение одной недели
после завершения старого тренировочного цикла
перед началом нового. Надо заметить, что советы
делать периодический перерыв в тренировках я
встречал и раньше, но переход к длительному
отдыху всегда мотивировался несколько иными
причинами — как правило, необходимостью в
психологической разгрузке или в заживлении
накопленных травм. Хейкок же прямо указал на то,
что полный отдых необходим для повышения
восприимчивости мышц к тренировке и назвал его
"стратегическим декондиционированием". Мой
интерес к HST объясняется ещё и тем, что данная
методика очень хорошо согласуется со всеми
сделанными мной ранее теоретическими выводами.
И хотя сам Хейкок привёл для обоснования
эффективности своей методики несколько иные
аргументы, я думаю, что HST стоит
проанализировать на основе предложенной мной
теории.
Суть методики HST заключается в следующем.
Из всего спектра существующих упражнений
выбираются 6-7 базовых, направленных на развитие
основных мышечных групп, далее все эти
упражнения выполняются в одном-двух рабочих
подходах на каждой тренировке три раза в неделю.
На первый взгляд может показаться, что столь
частая нагрузка на одни и те же мышцы — это
верный путь к перетренированности. Однако всё
дело в том, что тренировочный процесс в HST
разбивается на двухнедельные микроциклы (6
тренировочных занятий). В начале каждого
микроцикла спортсмен работает с заведомо
лёгкими весами, примерно 55-60% от
максимального веса, с которым возможно выполнить
конкретное упражнение в заданном количестве
повторений (не надо только путать с процентами от
максимума в единичном повторении). Вес снаряда
увеличивается от тренировки к тренировке на 5-10%
таким образом, что работа до "отказа" производится
лишь в последнюю тренировку микроцикла.
Соответственно, за пиком нагрузки в конце одного
микроцикла следует работа с относительно низкой
интенсивностью в начале следующего. Таким
образом, мышцы получают возможность
восстанавливаться в течение почти двух недель
вплоть до новой ударной нагрузки в конце нового
микроцикла (см. рис. 7). Прошу обратить
внимание на то, что в HST перерыв между
"отказными" тренировками составляет около двух
недель, равно как и в "Супертренинге" Менцера.
Ясно, что в отличие от редких тренировок
"Супертренинга" при частоте тренировок, принятых
в HST, мышцы адаптируются к работе значительно
быстрее, поэтому от микроцикла к микроциклу
воздействие ударных тренировок на сократительные
структуры мышц должно уменьшаться — но эта
проблема в HST решается постоянным повышением
средней интенсивности микроциклов. Так, в первом
микроцикле выполняется 15 повторений в подходе,
во втором — 10, в третьем — 5,
а в четвёртом, завершающем микроцикле обычные
повторения заменяются негативными, с весами
бо́льшими, чем в предыдущем микроцикле, то есть в
завершении макроцикла проводятся тренировки,
вызывающие максимальное количество микротравм.
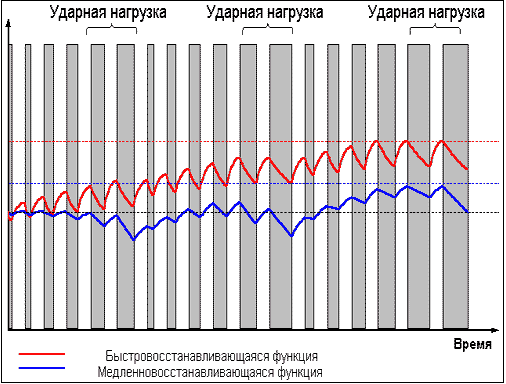 Рисунок 7
По завершении цикла негативных повторений мышцы
уже достаточно адаптированы к нагрузке, и
дальнейшие тренировки в таком состоянии будут
неэффективными для стимулирования роста мышечной
массы. По этой причине перед началом нового
макроцикла Хейкок вводит период "стратегического
декондиционирования" длительностью около недели
(я не исключаю возможность и более
продолжительного отдыха). За это время происходит
дезадаптация энергетики мышц, но количество
сократительных структур и, что главное, клеточных
ядер в мышцах остаётся на уровне, достигнутом в
ходе предыдущего макроцикла. После периода
"стратегического декондиционирования" тренировки
возобновляются по прежней схеме, начиная с
микроцикла на 15 повторений.
На мой взгляд, методика, предложенная Брайаном
Хейкоком, весьма удачно сочетает в себе
возможность полноценного восстановления
разрушенных тренировкой клеточных структур с
необходимостью регулярных и относительно частых
тренировок для повышения работоспособности мышц
и в то же время позволяет бороться с чрезмерной
адаптацией и снижением восприимчивости мышц к
нагрузке, что акцентирует воздействие тренировки
именно на мышечную гипертрофию.
Несмотря на тот факт, что об эффективности
периодической дезадаптации мышц для
стимулирования развития миофибриллярного
аппарата и о потребности в длительном
восстановлении клеточных структур после их
разрушения никто особенно не задумывался,
методика, учитывающая оба этих важных фактора,
применяется на практике достаточно давно. Я
веду речь о "силовом цикле" штангистов и
пауэрлифтёров. Настало время подробно разобрать
и эту методику тренировок, обладающую наивысшей
эффективностью для развития силы и окружённую
неким мистическим ореолом. Помню, первое время,
когда мне попадались на глаза расписанные по
неделям "процентовки", эти записи казались мне
сплошной кабалистикой, а между тем
варьирование нагрузки в течение "силового
цикла" подчиняется определённым законам и
преследует чёткие цели.
Из проделанного выше анализа следует, что сила
мышц зависит от четырёх основных факторов,
поддающихся тренировке: суммарного поперечного
сечения миофибрилл в мышцах; обеспеченности
миофибрилл энергией (концентрации АТФ
и скорости восполнения затрат АТФ —
мощности креатинкиназной реакции); способности нервной
системы к мгновенной и максимальной мобилизации
двигательных единиц; техники движения.
Понятно, что эффективная методика развития
силы должна положительно воздействовать на
все указанные компоненты.
Для увеличения совокупного поперечного сечения
миофибрилл посредством их сверхвосстановления
после микротравм нагрузка, преследующая эти цели,
должна быть достаточно длительной, дабы исчерпать
запасы креатинфосфата в ходе подхода (вспомните
условия возникновения микротравм). С другой
стороны, время под нагрузкой не должно быть
слишком большим, поскольку излишняя активизация
гликолиза нежелательна — ведь развитие
гликолитических возможностей мышц не сказывается
на максимальном результате в единичных
повторениях, но отвлекает на себя адаптационные
резервы организма. Отсюда следует, что
длительность тренирующей нагрузки должна находиться
у нижней границы эффективного воздействия на
миофибриллы. По этой причине в силовом цикле редко
работают более чем в пяти повторениях в подходе.
Такое количество повторений эффективно как для
стимулирования роста миофибрилл, так и для развития
креатинфосфатной мощности и ёмкости мышц. А для
этих целей, как я уже отмечал, требуется серия
истощающих креатинфосфат нагрузок, задаваемых через
интервал отдыха в несколько минут, достаточный для
суперкомпенсации запасов креатинфосфата после
истощения.
Но для развития креатинфосфатного потенциала мышц
требуется не менее двух-трёх тренировок в неделю.
С другой стороны, для обеспечения восстановления
мышц после микротравм требуется более длительный
отдых. Эта проблема решается уже знакомым способом
— циклированием нагрузки, причём в силовом
цикле прослеживается двойное циклирование.
Во-первых, из двух-трёх
тренировок в неделю на одну группу мышц только
одна является "тяжёлой" тренировкой, во время
которой и задаётся основная стимулирующая
нагрузка, остальные же тренировки являются
восстановительными и относятся к "лёгким".
Во-вторых, в тяжёлые дни нагрузка в
различные периоды "силового цикла" также меняется. Силовой
макроцикл разбивается на три основных микроцикла,
в ходе коих достигаются разные цели. Первый
микроцикл, длящийся несколько недель, называется
"втягивающим", его цель — подготовить
мышцы к предстоящей нагрузке путём развития их
креатинфосфатного потенциала и, тем самым,
обеспечения начально-необходимой
адаптированности энергетики мышц к предстоящей
работе (дабы последующие нагрузки не оказали излишне
разрушающее воздействие на мышцы).
Во время "втягивающего" микроцикла в основных
упражнениях, как правило, выполняется 5 подходов
по 5 повторений. Интенсивность в дни тяжёлых
тренировок повышается от недели к неделе
с 50-55% от единичного максимума в
начале и до 65% в конце микроцикла.
В лёгкие дни выполняется либо меньший объём движений
(количество поднятых штанг), либо используется
меньшая интенсивность, чем в дни тяжёлых
тренировок. Перерыва в одну неделю между
тяжёлыми тренировками при таких нагрузках
оказывается вполне достаточно для эффективного
восстановления мышц. За "втягивающим" микроциклом
следует несколько недель ударного микроцикла, в
течение которого в дни тяжёлых тренировок
интенсивность упражнений возрастает от 65%
до 75-80% от единичного максимума при
сохраняющемся количестве поднятых штанг.
В этот период мышцы получают максимальное
количество микротравм, и для полного восстановления
мышц недельного отдыха между тяжёлыми тренировками,
скорее всего, будет уже недостаточно. Поэтому
в течение "ударного" микроцикла происходит
накопление остаточного недовосстановления
мышц, и по достижении максимальной интенсивности
в конце микроцикла объём работы резко
сбрасывается путём уменьшения количества
повторений в подходе с пяти до двух-трёх.
Таким способом осуществляется переход к следующему
"восстановительному" микроциклу.
"Восстановительный" микроцикл длится ещё
несколько недель, в течение которых реализуется
отставленное после "ударного" микроцикла
сверхвосстановление мышц. Но роль последнего
микроцикла не сводится только к восстановлению
сократительного аппарата мышц после микротравм,
накопленных в "ударном" микроцикле, — в этот
период интенсивность тяжёлой тренировки
постепенно увеличивается от 80 до 100%,
что оказывает воздействие на способность мышц к
максимальной мобилизации двигательных единиц
и "приучает" сухожильные рецепторы к предельной
нагрузке. Примерно за неделю до соревнований
производится ещё одно снижение нагрузки, теперь
уже за счёт интенсивности (тренировочных весов),
что способствует максимальному восстановлению
нервной системы. В итоге "силовой цикл"
позволяет спортсмену к моменту соревнований
совместить пики развития основных тренируемых
функций и добиться максимального результата в
соревновательных движениях (см. рис. 8).
 Рисунок 8
(Колебания тренируемых функций после каждой
тренировки не показаны, приведены лишь
усреднённые значения)
При построении "силового цикла" важнейшей
задачей является совмещение во времени
максимумов развития всех основных компонентов
и подведение их именно к моменту соревнований,
поскольку постепенно достигнутая "форма" будет
утрачена ввиду перехода основных функций от
состояния суперкомпенсации к состоянию
утраченной суперкомпенсации. Снижение
креатинфосфатной ёмкости из-за уменьшения
нагрузки начинается уже во время
"восстановительного" микроцикла, что, кстати,
в совокупности с практикуемым после
соревнований недельным отдыхом выводит мышцы
из состояния адаптированности и повышает
эффективность воздействия тренировки на мышцы
во время следующего макроцикла.
Я описал обобщённую схему силового цикла,
каждый же конкретный цикл может отличаться от
остальных общей длительностью, длительностью
отдельных микроциклов внутри макроцикла,
количеством тренировочных занятий в неделю,
шагом интенсивности и рядом других параметров —
но общие принципы для всех "силовых циклов"
остаются практически неизменными.
На этом я, пожалуй, завершу анализ основных
тренировочных методик — хотя всё богатство и
разнообразие тренировочных "систем",
накопленных за годы развития "железного спорта",
не исчерпывается лишь теми методиками, которые
рассмотрены в данном тексте. Думаю, однако,
что теперь читатель сможет самостоятельно
оценить любую "новую" систему на предмет её
эффективности и соответствия поставленным целям.
Заключение
Во вступительном слове к данному тексту я обещал
доказать, что единая система в тренировках с
отягощениями всё же существует — несмотря на
все внешние различия и кажущиеся противоречия между
множеством практикуемых тренировочных методик.
Насколько мне это удалось сделать, придётся судить
читателю, но некоторые успехи очевидны, ибо я
выявил общие принципы тренировки, лежащие в
основе даже таких взаимоисключающих методик, как
высокоинтенсивный тренинг Майка Менцера и
объёмные тренировки "классического" бодибилдинга.
Боюсь, однако, что из-за относительно большого
объёма информации, обрушенной в тексте на читателя,
и специфики этой информации, выстроить в голове
изложенные мной сведения в систему удалось
далеко не всем. Поэтому сейчас, подводя итог данной
работы, я постараюсь сформулировать несколько
важнейших принципов, составляющих эту систему,
взяв за основу общие принципы спортивной
тренировки, приведённые Н.И.Волковым, слегка
уточнив их применительно к тренировкам с
отягощением и немного по-другому расставив
акценты.
1. Принцип суперкомпенсации
Следует понимать, что мышцы растут не на
тренировке, а во время отдыха после неё.
Тренировочная нагрузка вызывает изменения
внутренней среды мышц и организма в целом,
способствуя накоплению конечных продуктов
обмена веществ, растрачивая энергетические
ресурсы и разрушая активно работающие
клеточные структуры, что приводит к снижению
функции мышц и иных систем организма. По
завершении интенсивной работы организм
избавляется от продуктов метаболизма и
изношенных клеточных структур, одновременно
активизируются процессы восстановления
растраченных энергетических резервов и
синтез белка, необходимого для ремонта
повреждённых тканей. Интенсивные восстановительные
процессы, при условии достаточного по времени и
полноценного отдыха, приводят не просто к
восстановлению исходного состояния систем
организма, подвергшихся тренировочному
воздействию, но и обеспечивают превышение
функциональных возможностей этих систем над
дотренировочным уровнем (феномен суперкомпенсации).
В частности, восстановление разрушенных
тренировкой клеточных структур наряду с другими
факторами способствует гипертрофии мышечной
ткани. В соответствии с принципом
суперкомпенсации спортсмену следует уделять
внимание не только собственно тренировкам, но
и отдыху после них. Без суперкомпенсации
тренировки бессмысленны.
2. Принцип сверхотягощения и зависимость
"доза-эффект"
Зависимость между тренировочной нагрузкой и
достигаемым эффектом не является линейной. Слабая
нагрузка неспособна вызвать восстановительные
процессы такой интенсивности, чтобы обеспечить
заметную суперкомпенсацию тренируемых систем
организма. Существует порог величины
тренировочной нагрузки, ниже которого тренировка
не способна обеспечить сколько-нибудь заметное
воздействие на тренируемые функции; нагрузки ниже
этого порога являются неэффективными. Принцип
сверхотягощения заключается в том, что для
обеспечения суперкомпенсации тренируемых систем
организма необходимо, чтобы тренирующее
воздействие в достаточной степени нагружало и
побуждало их к развитию. Чем тяжелее тренировочная
нагрузка, тем выраженнее суперкомпенсация после
восстановления. Но адаптационные резервы
организма не беспредельны, и при нагрузках
определённой степени тяжести наблюдается уже
обратная ситуация: увеличение объёма и
интенсивности тренировки приводит к уменьшению
прироста результатов, и при сверхпредельных
нагрузках эффект от тренировки становится
отрицательным. В связи с этим очень важной задачей
является определение оптимальных тренировочных
нагрузок для каждого спортсмена.
3. Принцип положительного взаимодействия нагрузки
К сожалению, адаптационные изменения в организме
после единичной тренировки не закрепляются надолго.
При отсутствии повторной нагрузки на
соответствующую систему организма её, системы,
функция постепенно снижается до исходного
дотренировочного уровня, и фаза суперкомпенсации
постепенно сменяется фазой утраченной
суперкомпенсации. Поэтому редкие бессистемные
тренировочные воздействия неспособны вызвать
закрепление тренировочного эффекта и обеспечить
долговременную адаптацию организма. Слишком частые
нагрузки (до восстановления отягощённых
тренировкой функций) приводят к отрицательному
взаимодействию тренировочных эффектов и к
угнетению нагружаемых систем организма. И только
повторные нагрузки в состоянии суперкомпенсации
приводят к положительному взаимодействию
тренировочных эффектов и росту функциональных
возможностей спортсмена. Эффективная адаптация в
течение длительного периода тренировки становится
возможной только при условии положительного
взаимодействия между отдельными нагрузками.
4. Комплексное воздействие тренировки и принцип
специфичности
Общее воздействие тренировки на организм спортсмена
складывается из воздействия на отдельные функции и
системы. Тренировки с отягощением оказывают влияния
главным образом на эффективность нервно-мышечного
взаимодействия, алактатные и гликолитические
системы энергообеспечения мышц, сократительный
аппарат мышечных волокон, и в меньшей степени —
на аэробное энергообеспечение мышц и организма в
целом. Варьирование такими параметрами тренировки,
как длительность и интенсивность нагрузки,
позволяет управлять воздействием тренировки на
различные системы организма. Наиболее выраженные
адаптационные изменения происходят в системах,
нагружаемых в ходе тренировочного занятия в
наибольшей степени.
5. Гетерохронизм восстановительных процессов и
принцип циклирования нагрузки
Тренировка любой направленности одновременно
воздействует на целый ряд систем организма.
Согласно упомянутым ранее принципам
суперкомпенсации и положительного взаимодействия,
задачей спортсмена является обеспечение
восстановления всех нагружаемых в ходе тренировки
функций и систем. Различные системы и функции
имеют различное время восстановления
(гетерохронизм восстановительных процессов),
варьирующееся от нескольких минут и часов
(уровень АТФ и креатинфосфата в мышцах) до
нескольких дней и даже недель (восстановление
разрушенных клеточных структур).
Феномен гетерохронизма восстановительных
процессов не позволяет задать между тренировками
интервал отдыха, обеспечивающий положительное
взаимодействие срочных адаптационных изменений
во всех функциональных системах организма. При
любом раскладе нагрузок на момент новой
тренировки часть функциональных систем будет в
состоянии суперкомпенсации (положительное
взаимодействие нагрузок), часть в состоянии
утраченной компенсации (нейтральное
взаимодействие нагрузок), а часть систем ещё не
восстановится после прежних тренировок
(отрицательное взаимодействие нагрузок).
Исключить отрицательное взаимодействие нагрузок
можно только при относительно редких
тренировках, проводимых с частотой,
обеспечивающей достижение состояния
суперкомпенсации систем, имеющих самый
длительный период восстановления, либо при
более частых тренировках, исключив
сверхотягощение долговосстанавливающихся функций
(например, исключив разрушения клеточных
структур) — что не всегда возможно и не всегда
целесообразно.
При необходимости одновременного развития сразу
нескольких тренируемых функций, обладающих
свойством гетерохронизма, целесообразно проводить
тренировки с частотой, оптимальной для самой
быстровосстанавливающейся функции. Получить же
восстановление систем организма, требующих более
длительного отдыха, становится возможным только с
использованием метода циклирования нагрузки, то
есть варьирования объёмом и интенсивностью
тренировки. Метод циклирования нагрузки не
предусматривает суперкомпенсации тренируемых
функций к моменту каждого тренировочного занятия,
снижение функциональных возможностей спортсмена и
их сверхвосстановление достигается в определённые
периоды тренировочного процесса, называемые
микроциклами. Последовательное чередование
ударных и восстановительных микроциклов позволяет
получать суперкомпенсацию всех тренируемых
функций, несмотря на различия во времени,
требующемся на их восстановление.
6. Адаптация и принцип
прогрессивной нагрузки
По мере повышения тренированности организма
амплитуда возмущения его внутренней среды в ответ
на нагрузку уменьшается, соответственно снижается
и суперкомпенсация тренируемых систем организма.
Согласно принципу прогрессивной нагрузки, для
обеспечения адекватного воздействия на тренируемую
функцию тренировочная нагрузка должна планомерно
повышаться вслед за ростом тренированности
организма. С учётом рассмотренного ранее
принципа сверхотягощения и зависимости
"доза-эффект" следует понимать, что для
достижения оптимального тренировочного эффекта важна не
абсолютная, а относительная нагрузка — с
учётом текущего уровня тренированности спортсмена.
7. Адаптация и принцип
стратегического декондиционирования
Адаптационные изменения в организме со временем
могут повысить устойчивость ряда систем
организма к нагрузкам до такой степени, что
дальнейшее тренировочное воздействие на эти
системы, даже при условии повышения нагрузок, не
будет вызывать эффективную реакцию. В этой
ситуации оказывается целесообразным на
некоторое время прекратить нагрузки, вызвавшие
привыкание организма, и вернуться к тренировкам
адаптировавшихся систем только после их
частичного декондиционирования, когда
восприимчивость этих систем к нагрузке вновь
возрастёт. Декондиционирование
высокоадаптированных систем организма может быть
достигнуто либо переносом акцента тренировочного
воздействия на иные системы, либо полным отказом
от тренировок на некоторое время. Применительно
к тренировкам, направленным исключительно на
развитие мышечной массы за счёт сократительных
структур мышечных клеток, декондиционирование
систем энергообеспечения мышц (повышающее
микротравмирующий эффект тренировки) может носить
не только периодический характер, но и
реализовываться непрерывно по ходу тренировочного
процесса путём задания длительного интервала
отдыха между тренировками, — интервала, не
допускающего положительного суммирования
адаптационных изменений в энергетике мышц.
8. Предел адаптационных
возможностей организма и принцип
специализации
По мере приближения спортсмена к максимальным
возможностям своего организма суперкомпенсация
после тренировки снижается — даже при условии
достаточной нагрузки на тренируемые системы и
функции. Например, при максимальном развитии
мускулатуры спортсмена происходит замедление
дальнейшего прогресса не только по причине
снижения восприимчивости мышц и организма к
тренировке, но и вследствие приближения
возможностей систем организма, обеспечивающих
процессы восстановления мышечной ткани, к
естественным пределам. Ограничителями прогресса
становятся: гормональный фон спортсмена,
возможности пищеварительной системы по
обеспечению организма адекватным количеством
аминокислот, микроэлементов и другими
пластическими и энергетическими ресурсами.
Кроме того, увеличение диаметра мышечных волокон
создаёт естественный барьер на пути доставки
необходимых строительных и энергетических
элементов вглубь мышечных волокон. Все эти и
ряд других причин приводят к снижению
эффективности восстановления мышечной ткани после
повреждения, что неминуемо сказывается на
размере суперкомпенсационной прибавки.
По мере приближения спортсмена к пределу
адаптационных возможностей организма одним из
способов достижения дальнейшего прогресса, помимо
фармакологического вмешательства в естественные
процессы, становится отказ от попытки достижения
прогресса одновременно по всем возможным
направлениям, то есть специализация.
Специализация позволяет при достаточно сильном
воздействии на ограниченное количество
тренируемых систем организма снизить нагрузку на
общие системы жизнеобеспечения и, благодаря
перенаправлению всех ресурсов организма на
адаптацию в определённом направлении, добиться в
этом направлении максимального увеличения. В
силовых видах спорта специализация может
заключаться в отказе от попыток дальнейшего
развития мышечной ткани путём её
микротравмирования и в переключении только
на совершенствование нервно-мышечного
взаимодействия. Кроме того, можно
специализироваться на одном соревновательном
движении при поддержании результатов в остальных
движениях на достигнутом ранее уровне. В
бодибилдинге специализация может заключаться в
направлении тренировочного процесса на развитие
определённых мышечных групп на фоне
поддерживающей нагрузки на основные мышечные
массивы. Ряд методистов вообще считает
целесообразным снижение количества
выполняемых упражнений до минимума по мере
приближения спортсмена к пределу адаптационных
возможностей организма.
Уверен, что немало читателей смогли добраться до
конца моего текста лишь потому, что надеялись:
вот-вот я закончу изложение "скупой теории",
приступлю наконец к описанию "пышного зеленеющего
древа жизни" и, опираясь на открывшееся мне
"сокровенное" знание, сообщу магическую формулу
"научно обоснованной" методики тренировки —
когда, что и сколько им необходимо поднять для
осуществления заветной мечты (например, получения
бицепса окружностью 50 сантиметров). Боюсь их
разочаровать, но мне вспоминается чьё-то
мудрое изречение: "Если хочешь накормить человека, дай
ему не рыбу, а удочку". Я стремился не всучить
читателю очередную суперэффективную и "единственно
действенную" (ну как же иначе?) тренировочную
программу, а показать по возможности все пути,
ведущие к цели, и научить читателя самостоятельно
выбирать путь, наиболее эффективный и приемлемый
для него.
Опираясь на сформулированные выше общие принципы
тренировки и на конкретные сведения о различных
функциональных системах организма и о методах
воздействия на эти системы, приведённые
в третьей части данного текста, опытный спортсмен
в состоянии разработать оптимальную тренировочную
методику с учётом особенностей своего организма и
стоящих перед ним целей. Давать методические
советы профессионалам было бы с моей стороны
излишне самонадеянно — теория важна, но не
менее важно знание особенностей своего организма.
Вместе с тем я хотел дать и конкретные
практические рекомендации — но одним лишь
любителям, каковым я являюсь и сам.
Я уже как раз собрался сделать это в заключение
данного текста, но после нескольких безуспешных
попыток кратко изложить на бумаге мысли,
роящиеся у меня в голове, оставил сие
безнадёжное занятие. Разработка полноценных и
универсальных методических рекомендаций по
построению тренировочного процесса даже для
спортсмена-любителя требует сил и времени не
меньше, чем мне потребовалось на постижение и
изложение собственно теории. Возможно,
когда-нибудь позже я почувствую в себе силы и
решусь на это неблагодарное дело, но пока мне
не остаётся ничего другого, как только дать
совет из заголовка вышеприведённого текста:
"Думай!"
* * *
Литература:
- А.Д.Адо, М.А.Адо, В.И.Пыльницкий, Г.В.Порядина, Ю.А.Владимирова "Патологическая физиология" Москва (2001)
- Т.Т.Берёзов, Б.Ф.Коровкин "Биологическая химия" Москва "Медицина" 1998 г.
- П.Г.Богач, В.Н.Дубонос, В.Л.Зима, В.М.Данилова "Конформационные изменения миозина при различных рН раствора" сборник "Биофизика и биохимия мышечного сокращения" Москва 1976 г.
- М.Е.Валиулина "Роль нейротрофического контроля и гуморальной регуляции синтеза миозинов скелетных мышц" Казань 1996 г.
- Н.И.Волков, Э.Н.Нессен, А.А.Осипенко, С.Н.Корсун "Биохимия мышечной деятельности" Киев (2000)
- А.В.Володина "Посттравматическая регенерация скелетных мышц" Москва 1995 г.
- Н.А.Габелова, К.С.Алейникова "Процесс расслабления и пластификации миофибрилл" (сборник "Биофизика мышечного сокращения") Москва 1966 г.
- Владимир Гончаров "Логика тренинга"
- В.С.Гурфинкель, Я.М.Коц, М.Л.Шик "Регуляция позы человека" Москва "Наука" 1965 г.
- В.С.Гурфинкель, О.С.Левин "Скелетная мышца структура и функция" Москва "Наука" 1985 г.
- В.В.Дынник "Внутриклеточные механизмы контроля скоростей синтеза и гидролиза АТФ в мышцах" сборник научных трудов "Механизмы контроля мышечной деятельности" Ленинград "Наука" 1985 г.
- В.И.Дещеревский "Экспериментальные основы и постулаты кинетической теории мышечного сокращения" сборник научных трудов "Механизмы мышечного сокращения" Москва "Наука" 1972 г.
- Н.П.Дещеревская "О возможной роли двух головок миозина в мышечном сокращении" Санкт Петербург (1991)
- Р.П.Женевская "Нервно-трофическая регуляция пластической активности мышечной ткани" Москва 1974 г.
- Е.К.Жуков "Очерки по нервно-мышечной физиологии" Ленинград 1969 г.
- М.М.Заалишвили "Метаболизм мышечной клетки и некоторые вопросы энергетики мышечного сокращения" в сборнике "Структурные основы и регуляция биологической подвижности" Москва 1980 г.
- И.И.Иванов "Об использовании энергии АТФ при мышечном сокращении" сборник научных трудов "Биофизика мышечного сокращения" Москва "Наука" 1966 г.
- А.А.Климов Р.К.Данилов "Миосателлитоциты" Архангельск 1981 г.
- В.Ф.Кондиаленко, Ю.Д.Сергеев, В.В.Иваницкая "Электронно-микроскопическое исследование проявлений гиперплазии мышечных волокон скелетных мышц спортсменов" Архангельск 1981 г.
- Я.М.Коц "Физиология мышечной деятельности" Москва 1982 г.
- А.Ф.Краснова "Биохимические изменения в мышцах и крови при мышечной деятельности силового характера" Ленинград 1960 г.
- Н.А.Кубасова "Исследование механических и структурных свойств миозинового мостика" (2000)
- В.П.Куликов, В.И.Киселёв "Потребность в двигательной активности" Новосибирск "Наука" (1998)
- В.В.Леднёв "Некоторые аспекты регуляции мышечного сокращения" сборник "Структурные основы и регуляция биологической подвижности" Москва 1980 г.
- Л.Г.Магазаник, Г.А.Наследов "Развитие сократительной функции мышц двигательного аппарата" Ленинград 1974 г.
- А.Дж.Мак-Комас "Скелетные мышцы" Киев "Олимпийская литература" (2001)
- С.Мак-Роберт "Думай! Бодибилдинг без стероидов" "Медиа спорт" (1998)
- М.Менцер "Супертренинг" "Медиа спорт" (1998)
- В.И.Морозов "Биохимические механизмы участия лейкоцитов в метаболическом ответе скелетных мышц на физическую нагрузку" Санкт-Петербург 1997 г.
- Б.Ф.Поглазов "Миозин и биологическая подвижность" Москва 1982 г.
- Н.И.Разумовская "Роль нервной системы в регуляции синтеза мышечных белков" сборник "Нервный контроль структурно-функциональной организации мышц" Ленинград 1980 г.
- Д.Ю.Романовский "Влияние инкубационной среды скелетных мышц на утомление нервно мышечного препарата и на освобождение медиатора в нервно-мышечном синапсе лягушки" Санкт-Петербург 1996 г.
- А.Б.Рубин "Биофизика" в 2-х томах Москва 2000 г.
- В.А.Сакс, В.В.Куприянов "Внутриклеточный транспорт энергии" сборник "Биохимия и биофизика мышц" Москва 1989 г.
- В.А.Сакс, Э.К.Сеппет, В.В.Куприянов, В.Н.Смирнов "Механизмы энергообеспечения мышечного сокращения" в сборнике "Структурные основы и регуляция биологической подвижности" Москва 1980 г.
- М.Сингер, П.Берг "Гены и геномы" Москва "Мир" 1998 г.
- С.Л.Соков, Л.П.Соков "Информационное моделирование адаптационных синдромов травматических стресс-ситуаций" Вестник Российского университета дружбы народов Серия "Медицина" 1999 № 1. С. 91-99
- Х.Тюннеманн, Ю.Хартманн "Современная силовая тренировка. Теория и практика" Берлин 1988 г.
- Э.Г.Улумбеков, Н.П.Ревзяков "Нейротрофический контроль фазных мышечных волокон" сборник "Нервный контроль структурно-функциональной организации мышц" Ленинград 1980 г.
- Э.Г.Улумбеков, Ю.А.Челышев "Гистология, введение в патологию", Москва "ГЕОТАР Медицина" 1998 г.
- Г.М.Харгивс "Метаболизм в процессе физической деятельности" Киев "Олимпийская литература" (2001)
- Фредерик К.Хэтфилд "Всестороннее руководство по развитию силы" Новый Орлеан, 1983 г.
- М.Хосни "Биоэнергетика повторной мышечной работы и эффективность интервальной тренировки в спорте" Москва 1995 г.
- Б.С.Шекман "Влияние тренировки на композицию мышц, размер и окислительный потенциал мышечных волокон у человека" Москва 1990 г.
- Р.Шмидт, Г.Тевс "Физиология человека" в 3-х томах, Москва "Мир" 1996 г.
- М.Д.Штерлинг, Е.Е.Филютина, И.И.Бугаева, О.Л.Гребнёва, Н.А.Плотникова "Скелетная мышца. Структурные аспекты адаптации" Новосибирск 1991 г.
- И.Т.Штракфельд, И.Е.Москаленко "Изменения двигательного аппарата мышц при денервации" сборник "Нервный контроль структурно-функциональной организации мышц" Ленинград 1980 г.
- Cabric D.L., Daniels J., Evanes W. Et al "Skeletal muscle enzymes and fiber composition in male and female track athletes" J Appt Physiol. — 1976
- Gonyea W, Ericson GC, Bonde-Petersen F "Skeletal muscle fiber splitting induced by weight-lifting exercise in cats"
- Haycock B "The hypertrophy specific training", www.thinkmuscle.com
- Hatfield F "The simplicity of periodicity", www.drsquat.com
- Larsson L & Tesch PA (1986) "Motor unit fiber density in extremely hypertriphied skeletal muscles in man" Eur J Appl Physiol 55:1130-1136
- Yamada S, Buffinger N, Dimario J & Strohman R (1989) "Fibroplast growth factor is stored in fiber extracellular matrix and plays a role in regulating muscle hypertrophy" Med Sci Sports Exerc 21(5): S173-s180
***
Как Вадим Протасенко в конце концов выяснил,
мышечные волокна получают наибольшее число
микротравм действительно во время уступающих
движений даже с относительно небольшой
нагрузкой — то есть это отнюдь не
заблуждение, а достоверный факт — Сост.

[на главную страницу]
Архив переписки
Форум
| |